


Transport des lipides
Lipoprotéines : récepteurs
Récepteurs LDLR : LRP4 (Megf7) et système nerveux périphérique
- Biochimie
- Chimie organique
- Bioénergétique
- Composition de la matière vivante
- Composés organiques
- Protides
- Acides nucléiques
- Glucides
- Lipides
- Vue d'ensemble des lipides
- Acides gras
- Classification des lipides
- Cholestérol
- Glycérides
- Trafic non vésiculaire des lipides et LTP
- Digestion et absorption entérocytaire des lipides
- Lipoprotéines
- Gouttelettes lipidiques
- Coenzymes
- Hormones
- Composés inorganiques
- Composés organiques
- Transport membranaire
- Moteurs moléculaires
- Voies de signalisation

LRP4 (Megf7), à part son rôle lié à l'agrine dans le développement des jonctions neuromusculaires (NMJ), interviendrait dans d'autres mécanismes antérieurs à la différenciation post-synaptique des NMJ.
LRP4 et système nerveux périphérique
Tout au long du développement de la NMJ et contrairement à son rôle dans le regroupement synaptique en tant que récepteur de l'agrine, LRP4 peut également fonctionner au niveau de la présynapse, indépendamment de l'agrine, et de manière autonome non cellulaire ( LRP4/agrine dans les NMJ).
LRP4/agrine dans les NMJ).
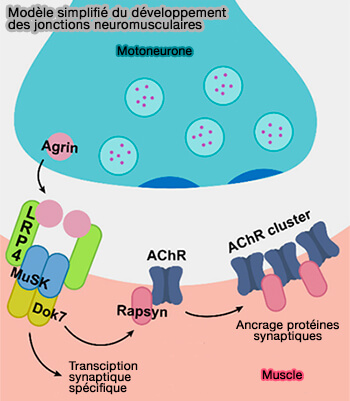
(Figure : vetopsy.fr d'après DePew et Mosca)
Au niveau de la jonction neuromusculaire des mammifères, LRP4 instruit plusieurs étapes de développement pour réguler la différenciation pré- et post-synaptique qui pourraient être les suivantes (Conservation and Innovation: Versatile Roles for LRP4 in Nervous System Development 2021).
1. Avant l'arrivée du motoneurone, LRP4 est nécessaire pour la pré-structuration ou pré-modelage (prepatterning) des récepteurs de l'acétylcholine (AChR) dans la région synaptique future (Patterning of Muscle Acetylcholine Receptor Gene Expression in the Absence of Motor Innervation 2001)..
2. À l'arrivée du motoneurone, LRP4 est nécessaire comme signal d'arrêt pour que le cône de croissance arrête tout déplacement le long de la fibre musculaire, i.e. rôle essentiel pour l'initiation du contact adhésif entre le motoneurone et le muscle (Distinct Roles of Different Presynaptic and Postsynaptic NCAM Isoforms in Early Motoneuron–Myotube Interactions Required for Functional Synapse Formation 2018).
3. Une fois le contact synaptique effectué, LRP4 ordonne alors la différenciation présynaptique des motoneurones (Distinct Roles of Muscle and Motoneuron LRP4 in Neuromuscular Junction Formation 2012).
En l'absence de LRP4, les motoneurones sont incapables de regrouper les protéines de la zone active et les vésicules synaptiques (Lrp4 Is A Retrograde Signal For Presynaptic Differentiation At Neuromuscular Synapse 2012).
LRP4 fonctionnerait comme un signal rétrograde pour instruire la différenciation présynaptique en interagissant avec un récepteur encore non identifié sur les axones moteurs, indépendamment de la liaison à l'agrine et de l'activation de MuSK.
- Ce signal serait fourni par le domaine extracellaire de LRP4 qui aurait été clivé au niveau des NMJ et serait suffisant pour instruire la formation de NMJ sans être ancré à la membrane cellulaire (The extracellular region of Lrp4 is sufficient to mediate neuromuscular synapse formation 2011).
- Le mécanisme protéolytique serait probablement du à l'activité de MMP (Matrix MetalloProteinase) (Neuronal MT1-MMP mediates ECM clearance and Lrp4 cleavage for agrin deposition and signaling in presynaptic development 2020).

(Figure : vetopsy.fr d'après DePew et Mosca)
4. LRP4 signale également la différenciation postsynaptique en se liant à l'agrine et en stimulant l'activation de MuSK comme vu dans le chapitre précédent.
LRP4 et signalisation Wnt
La voie agrine n'est pas la seule à provoquer le groupement des AChR, ni le seul mécanisme de régulation sous-jacent à la formation de NMJ (Neuromuscular Junction Formation, Aging, and Disorders 2018).
- Les étapes essentielles de la pré-structuration des AChR fonctionnent de manière indépendante de l'agrine ( cf. plus haut).
- Elles reposent probablement sur des mécanismes alternatifs tels que " l'auto-activation " LRP4/MuSK et la voie de signalisation Wnt (Diverging roles for Lrp4 and Wnt signaling in neuromuscular synapse development during evolution 2016).
1. Les voies Wnt régulent la différenciation et la spécialisation cellulaires en impliquent la liaison des molécules Wnt aux récepteurs Frizzled transmembranaires, via des mécanismes de signalisation distincts (WNT Signaling in Disease 2019) :
- la stabilisation " canonique " de la β-caténine régulant l'expression des gènes qui nécessite une interaction avec LRP5/6 ( LRP5/6 et voie Wnt).
- les voies " non canoniques " de polarité cellulaire planaire (PCP) et Ca++/Calmoduline qui influencent le réarrangement du cytosquelette.
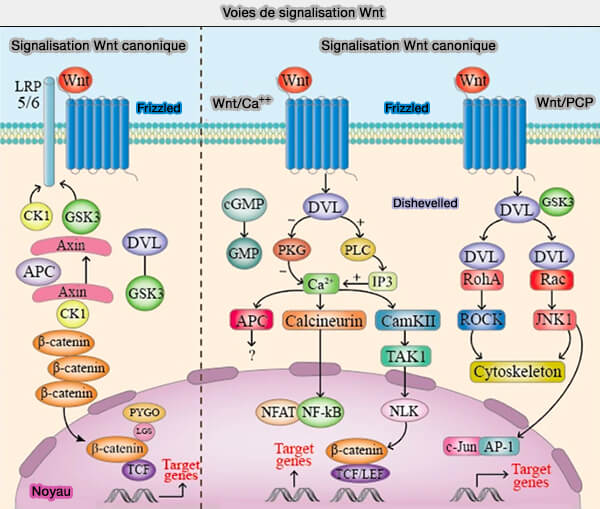
(Figure : vetopsy.fr d'après Zhao et coll)
2. Les voies Wnt semblent exister dans une sorte d'équilibre dynamique canonique/non canonique (Dissecting the Extracellular Complexity of Neuromuscular Junction Organizers 2020).
a. La signalisation canonique Wnt semble être nécessaire pour le pré-structuration/regroupement aneural de l'AChR (Wnt proteins contribute to neuromuscular junction formation through distinct signaling pathways 2017).
Toutefois, elle régule négativement le regroupement des AChR par la rapsyne (Wnt/β-Catenin Signaling Suppresses Rapsyn Expression and Inhibits Acetylcholine Receptor Clustering at the Neuromuscular Junction 2008).
b. À l'inverse, favoriser la signalisation non canonique Wnt via DKK1 (Dickkopf1), entraîne une pré-structuration altérée ainsi qu'une ramification aberrante des motoneurones et de la formation des NMJ.
c. Les modèles de souris à gain de fonction ou perte de fonction de la β-caténine affichent une formation de NMJ perturbée de manière similaire (β-Catenin stabilization in skeletal muscles, but not in motor neurons, leads to aberrant motor innervation of the muscle during neuromuscular development in mice 2012).

(Figure : vetopsy.fr d'après Guarino et coll)
3. Cependant, le mode de signalisation Wnt à la jonction neuromusculaire est encore obscur et fait débat avec deux théories.
a. La théorie la plus plausible est celle de la médiation via la voie LRP4/MuSK.
- Des Wnt se lient se lier directement à MuSK, i.e. Wnt4, Wnt11, Wnt9a, et LRP4, i.e. Wnt9a, Wnt11 (Wnt proteins regulate acetylcholine receptor clustering in muscle cells 2012).
- MuSK interagit avec le médiateur Wnt Disheveled (Dsh) intracellulaire, qui se situe au carrefour entre les voies canoniques et non canoniques Wnt (Regulation of AChR Clustering by Dishevelled Interacting with MuSK and PAK1 2002).
- Les inhibiteurs de la voie canonique Wnt tels que DKK1 (Dickkopf1), la SOST (sclérotine) et les protéines morphogéniques osseuses (BMP) se lient à LRP4, fournissant ainsi une couche de modulation supplémentaire possible (LRP4 in neuromuscular junction and bone development and diseases 2015).

Le modèle possible du mécanisme est décrit dans : Dissecting the Extracellular Complexity of Neuromuscular Junction Organizers (2020).
b. La théorie la moins plausible est celle qui inclue la signalisation via Frizzled canonique/LRP5/6, la médiation par LRP4 (nécessaire pour la pré-structuration) via un récepteur non identifié.
Retour aux LDLR
BiochimieChimie organiqueBioénergétiqueProtidesGlucidesLipidesAcides grasPhospholipidesPhosphoinositidesCholestérolAcides biliairesGlycéridesDigestion et absorption entérocytaire des lipidesLipoprotéinesApolipoprotéinesTrafic des lipoprotéinesRécepteurs des lipoprotéinesGouttelettes lipidiques (LD)EnzymesCoenzymesVitaminesHormonesComposés inorganiquesTransport membranaireMoteurs moléculairesVoies de signalisation