


Nucléotides
Ribonucléotides
Régulation de la biosynthèse
- Biochimie
- Transport membranaire
- Moteurs moléculaires
- Voies de signalisation

La biosynthèse des nucléotides puriques et pyrimidiques est régulée à plusieurs niveaux enzymatiques afin de contrôler la production totale de nucléotides et de maintenir l’équilibre entre leurs différentes formes.
1. La biosynthèse de novo des ribonucléotides puriques et pyrimidiques est soumise à plusieurs mécanismes de régulation permettant d’adapter la production de nucléotides aux besoins métaboliques de la cellule.
- Ces mécanismes contrôlent à la fois le flux global de synthèse et l’équilibre entre les différentes familles de nucléotides.
- Ils s’exercent principalement au niveau des premières étapes de chaque voie biosynthétique, ainsi qu’au niveau des points de bifurcation métabolique qui déterminent la formation des principaux nucléotides puriques et pyrimidiques.
2. Cette régulation assure le maintien d’un pool équilibré de nucléotides indispensable à la synthèse des acides nucléiques et au bon fonctionnement du métabolisme cellulaire.
Régulation de la biosynthèse des nucléotides puriques
La synthèse de novo des purines :
- commence avec le 5-PRPP (phosphoribosylpyrophosphate) et produit l'IMP,
- l'IMP est le précurseur commun de l’AMP et du GMP.
Régulation de la synthèse totale des purines
La biosynthèse de novo des purines est soumise à un contrôle global qui permet d’éviter une production excessive de nucléotides puriques lorsque ceux-ci sont déjà présents en quantité suffisante dans la cellule.
Ce contrôle s’exerce principalement au niveau des premières étapes de la voie.
Premier contrôle
Le premier contrôle s'effectue au niveau de la synthèse du 5-PRPP (5-phosphoribosylpyrophosphate) par la ribose-phosphate diphosphokinase.
Cette enzyme est inhibée par plusieurs nucléotides, notamment l'ADP et le GDP, ce qui permet de limiter la formation de PRPP lorsque les pools intracellulaires de nucléotides sont suffisants.
Ce mécanisme constitue un contrôle global en amont de la biosynthèse des nucléotides, le 5-PRPP étant le substrat initial des voies de synthèse des purines ( étape princeps) et des pyrimidines (cinquième réaction).
étape princeps) et des pyrimidines (cinquième réaction).
Deuxième contrôle
Le deuxième contrôle s'effectue au niveau de la transformation du 5-PRPP en 5-phosphoribosylamine (PRA), réaction 1 de la formation de l'IMP.
1. L'amidophosphoribosyltransférase (ATase), appelée aussi glutamine phosphoribosylpyrophosphate amidotransférase ou Glutamine PRPP AmidoTransférase (GPAT), EC 2.4.2.14, est un homotétramère qui comporte deux domaines structuraux sur chaque sous-unité (Crystal structure of glutamine phosphoribosylpyrophosphate amidotransferase from Escherichia coli 1998 et Glutamine PRPP amidotransferase: snapshots of an enzyme in action 1998).
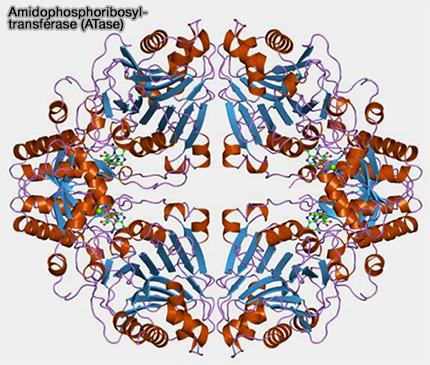
(Figure : vetopsy.fr)
a. Le domaine N-terminal (domaine glutaminase) hydrolyse l'amide de la glutamine :
$\ce{glutamine + H2O}$ $\leftrightharpoons$ $\ce{NH3 + glutamate}$
b. Le domaine C-terminal (domaine PRTase), site actif du domaine phosphoribosyltransférase se lie au PRPP et utilise le NH3 pour la synthèse du PRA :
$\ce{PRPP + NH3}$ $\leftrightharpoons$ $\ce{PRA + PPi}$.
2. Le domaine C-terminal possède deux sites inhibiteurs de liaison aux nucléotides qui chevauchent le domaine PRTase :
- le site C qui chevauche le site pour le ribose-5-phosphate de PRPP,
- le site A allostérique chevauche le site pour le pyrophosphate de PRPP.
a. Lorsque les nucléotides puriques finaux, i.e. AMP, ADP, GMP ou GDP, s'accumulent dans la cellule, ils peuvent se fixer sur ces sites inhibiteurs.
b. Cette fixation empêche la fermeture de la boucle flexible du domaine PRTase, étape nécessaire à la liaison du 5-PRPP au site actif, et bloque ainsi la catalyse.
- Ce blocage résulte d'interactions électrostatiques entre les nucléotides inhibiteurs et la base de la boucle flexible du domaine PRTase, impliquant notamment les résidus Lys326 et Arg328 (E. coli GPATase).
- Ces résidus interagissent normalement avec le substrat PRPP, mais leur interaction avec les nucléotides inhibiteurs empêche la fixation correcte du substrat.
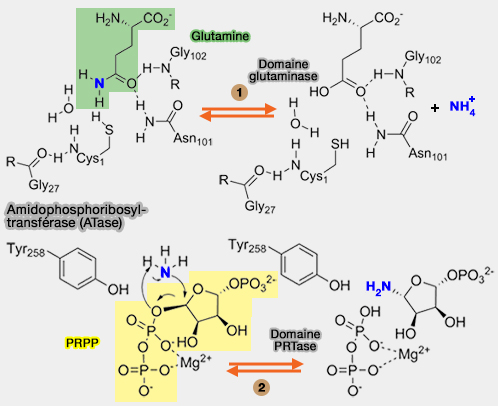
(Figure : vetopsy.fr)
3. Les deux sites inhibiteurs A et C présentent en outre une forte synergie fonctionnelle, car chacun d’eux possède une préférence de liaison pour des nucléotides puriques différents.
a. En effet, selon les organismes :
- le site A fixe préférentiellement des nucléotides dérivés de l’adénine, i.e. l'AMP ou l'ADP,
- le site C fixe plutôt des nucléotides dérivés de la guanine, i.e. le GMP ou le GDP.
b. Lorsque seul l’un de ces nucléotides est présent en excès, l’inhibition de l’enzyme reste partielle et la synthèse de novo peut se poursuivre.
L’IMP produit est alors orienté préférentiellement vers la formation du nucléotide qui manque, ce qui contribue à rétablir l’équilibre entre dérivés de l’adénine et de la guanine (maintien de l’équilibre AMP/GMP).
b. En revanche, lorsque les nucléotides dérivés de l’adénine et de la guanine sont tous deux abondants, ils occupent simultanément les deux sites inhibiteurs, ce qui provoque une inhibition synergique de l’enzyme et bloque efficacement la synthèse de l’IMP.

Le résultat physiologique est la régulation fine de la première étape de la biosynthèse des purines grâce à l'inhibition synergique par la combinaison de ces nucléotides.
Maintien de l’équilibre AMP/GMP
Au-delà du contrôle global de la synthèse des purines, la cellule doit également maintenir un équilibre entre les deux nucléotides puriques principaux, l’AMP et le GMP.
1. Tout d'abord, on assiste à une rétroinhibition spécifique :
- l'AMP inhibe la conversion de l'IMP en adénylosuccinate, précurseur immédiat de l’AMP dans la voie de biosynthèse,
- le GMP inhibe la conversion de l'IMP en XMP, précurseur immédiat de l’AMP dans la voie de biosynthèse.
2. Par ailleurs, il existe une régulation croisée énergétique :
- la conversion de l'IMP en AMP nécessite du GTP,
- la conversion de l'IMP en GMP nécessite de l'ATP.
Cette régulation croisée permet d’équilibrer la synthèse des deux nucléotides : un excès de l’un des nucléotides stimule indirectement la synthèse de l’autre, en fournissant l’énergie nécessaire à sa formation.
Régulation de la biosynthèse des nucléotides pyrimidiques
La biosynthèse des ribonucléotides pyrimidiques est soumise à plusieurs niveaux de régulation qui permettent d’adapter la production de nucléotides aux besoins cellulaires et de maintenir l’équilibre entre nucléotides puriques et pyrimidiques.
Régulation de l’entrée de la voie pyrimidique
Le premier contrôle s'effectue au niveau de la formation du carbamyl-phosphate (réaction 1 de la formation de l'UMP) par la carbamyl-phosphate synthase II (CPSase II) qui est :
1. activée par :
- l'ATP, signal d’un état énergétique élevé de la cellule,
- le PRPP, indicateur d’un besoin en nucléotides,
2. inhibée par :
- l'UMP, produit final de la voie (rétro-inhibition),
- l'UTP, qui assure un contrôle fin sur le flux pyrimidique.
Régulation au niveau de l’aspartate transcarbamylase (ATCase)
1. Un second contrôle intervient lors de la formation du carbamyl-aspartate (réaction 2 de la formation de l'UMP), catalysée par l’aspartate transcarbamylase (ATCase).
Cette enzyme constitue un point majeur de régulation de la voie pyrimidique.
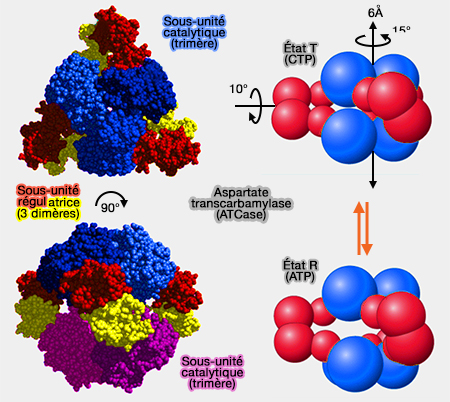
(Figure : vetopsy.fr d'après esilrch1.esi.umontreal.ca)
2. L’ATCase est composée de :
- 6 sous-unités catalytiques formant 2 trimères catalytiques,
- 6 sous-unités régulatrices formant 3 dimères régulateurs.
a. L’activité de l’ATCase dépend d’une transition conformationnelle entre deux états :
- la forme T, i.e. tendue, à faible affinité pour le substrat, stabilisée par les nucléotides pyrimidiques, notamment le CTP et surtout le dCTP, mais aussi plus faiblement le dTTP et l’UTP, ce qui inhibe l’enzyme,
Le dCTP exerce l’inhibition la plus forte car il reflète directement l’abondance des désoxyribonucléotides utilisés pour la synthèse de l’ADN.
- la forme R, i.e. relâchée, à forte affinité pour le substrat, favorisée par les nucléotides puriques, notamment l’ATP et le dATP, ce qui active l’activité enzymatique.
b. La transition T ➞ R s’accompagne d’un changement structural mesurable.
La distance entre les trimères catalytiques et les sous-unités régulatrices passe d’environ 12 Å à 4 Å.
Cette régulation allostérique permet d’équilibrer la synthèse des nucléotides pyrimidiques avec celle des nucléotides puriques.
Régulation de la formation du CTP
La dernière étape de la biosynthèse des ribonucléotides pyrimidiques correspond à la conversion de l’UTP en CTP catalysée par la CTP synthase ( formation du CTP).
$\ce{ATP + UTP + NH3}$ $\leftrightharpoons$ $\ce{ADP + phosphate + CTP}$
La CTP synthase contrôle ainsi l’équilibre entre UTP et CTP.
1. La CTP synthase est :
- inhibée par le CTP, son produit final, qui agit comme effecteur allostérique négatif et empêche une surproduction de CTP
- activée par le GTP, qui favorise la conformation active de l’enzyme.
Le GTP facilite notamment la transmission de l’ammoniac (NH3) généré par l’hydrolyse de la glutamine vers le site de l’UTP, améliorant l’efficacité de l’amination.
2. Cette régulation permet de maintenir un équilibre adéquat entre UTP et CTP, nécessaire à :
- la synthèse d’ARN,
- la biosynthèse des phospholipides membranaires,
- le métabolisme cellulaire.
3. La régulation de la CTP synthase s’intègre dans un réseau global de contrôle métabolique qui coordonne la biosynthèse des nucléotides puriques et pyrimidiques pour répondre aux besoins physiologiques.
L'interconversion des nucléotides est étudiée dans un chapitre spécifique.
Retour à la biosynthèse des nucléotides
BiochimieChimie organiqueBioénergétiqueAcides nucléiquesNucléotidesBases azotéesPentosesDifférents nucléosidesDifférents nucléotidesBiosynthèse des nucléotidesVoies des sauvetageCatabolisme des nucléotidesADNARNChromatineNucléosomesHistonesChromosomesProtidesGlucidesLipidesEnzymesCoenzymesVitaminesHormonesComposés inorganiquesTransport membranaireMoteurs moléculairesVoies de signalisation