La séparation de phase liquide-liquide (LLPS) est un processus physico-chimique au cours duquel un mélange homogène de macromolécules en solution aqueuse se démixe spontanément en deux phases liquides coexistantes, une phase diluée et une phase dense enrichie en composants spécifiques.
C'est par exemple la séparation de l'huile dans l'eau.
Séparation de phase liquide-liquide (LLPS)
Vue d'ensemble
En biologie cellulaire, il s’agit d’un processus par lequel des macromolécules se démixent d’un mélange aqueux homogène pour former une phase dense distincte, communément appelée condensat biomoléculaire ou organite sans membrane (Liquid phase condensation in cell physiology and disease 2017).
Séparation huile/eau
(Figure : vetopsy.fr d'après Dr Detailling)
En atteignant la concentration de saturation dans une solution, i.e. concentration dite critique, ces biomolécules s’assemblent spontanément en une phase dense dépourvue de membrane, pouvant néanmoins impliquer des composants organisateurs multivalents jouant un rôle nucléateur.
Condensats biomoléculaires organisateurs en biologie neuronale et synaptique
(Figure : vetopsy.fr d'après Sansevrino et coll)
3. Une telle phase dense peut enrichir sélectivement certaines molécules et en exclure d’autres en fonction de leur valence d’interaction, de leur affinité et de leurs propriétés physico-chimiques (Compositional Control of Phase-Separated Cellular Bodies 2016).
Les condensats biomoléculaires présentent plusieurs propriétés physiques caractéristiques des phases liquides concentrées, parmi lesquelles :
la fusion possible de deux condensats voisins qui forment une gouttelette unique,
une tendance à adopter une forme sphérique résultant de la tension de surface,
4. Les condensats ne sont toutefois pas des structures statiques et peuvent évoluer au cours du temps.
Leur état peut progressivement passer d’une phase liquide dynamique à des états plus viscoélastiques ou gélifiés, phénomène parfois qualifié de " vieillissement " des condensats et impliqué notamment dans certaines maladies neurodégénératives comme la sclérose latérale amyotrophique (SLA) ou certaines formes de démence frontotemporale, associées à l’agrégation de protéines telles que FUS ou TDP-43 ( pathologies des corps nucléaires).
L’assemblage et la dissolution des condensats sont également régulés par des modifications post-traductionnelles des protéines impliquées.
Des modifications telles que la phosphorylation, la SUMOylation, l’ubiquitination ou la méthylation des arginines peuvent modifier la valence d’interaction des protéines et moduler ainsi leur capacité à entrer ou sortir d’un condensat.
Extension du LLPS aux organites
1. Les mécanismes de séparation de phases sont indépendants de l’échelle spatiale et concernent principalement des assemblages macromoléculaires dynamiques ; ils ne transforment pas un organite entier en phase séparée, mais peuvent organiser localement certains de ses composants en microdomaines condensés.
a. Les corps nucléaires constituent des exemples majeurs de tels assemblages.
Modèles de condensation par la synapsine
(Figure : vetopsy.fr d'après Zhang et coll)
c. Des mécanismes analogues interviennent également dans l’organisation présynaptique et la dynamique des vésicules synaptiques (VS), qui font l’objet d’un développement spécifique dans la page consacrée aux synapses.
2. De plus, l'activité biologique de certaines biomolécules dépend de leur capacité à former des condensats.
Autrement dit, leur fonction ne repose pas uniquement sur leur présence, mais sur leur capacité à se concentrer transitoirement dans un espace restreint afin d’augmenter localement les interactions moléculaires.
a. Par exemple, dans les neurones, la formation et le transport des granules d'ARN sont étroitement couplés à leur capacité à former des condensats biomoléculaires.
Ces granules correspondent à des regroupements dynamiques d’ARN et de protéines qui se comportent comme des microgouttelettes liquides capables de se déplacer dans le cytoplasme.
Plus ces condensats présentent des propriétés liquides adaptées, plus ils peuvent interagir de manière transitoire avec des organites moteurs comme les lysosomes, facilitant ainsi leur transport sur de longues distances dans l’axone.
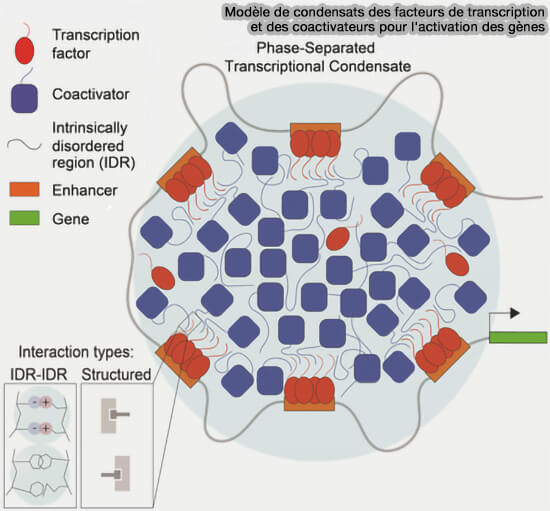
Modèle de condensats des facteurs de transcription
(Figure : vetopsy.fr d'après Boijja et coll)
c. Certains facteurs de transcription et coactivateurs possédant des régions intrinsèquement désordonnées peuvent former des condensats au niveau des enhancers ou des super-enhancers, concentrant localement l’ARN polymérase II et les cofacteurs nécessaires à l’initiation transcriptionnelle.
1. De nombreuses protéines impliquées dans la formation de condensats contiennent une région intrinsèquement désordonnée (IDR ou Intrinsic Disordered Region).
Les IDR sont des suites d'acides aminés qui ne se replient pas dans une structure secondaire ou tertiaire stable.
des protéines des granules de stress cytoplasmiques telles que G3BP1, acteur central de leur nucléation, et FUS ou TDP-43 impliquées dans des condensats ribonucléoprotéiques dynamiques,
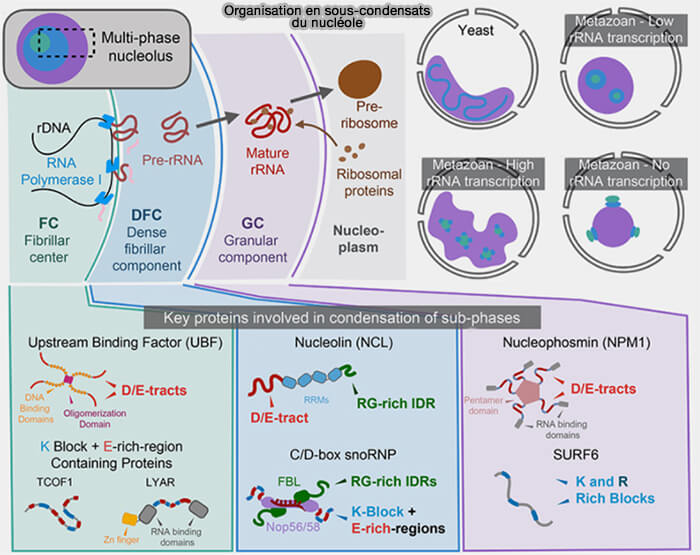
Organisation en sous-condensats du nucléole
(Figure : vetopsy.fr d'après King et coll)
3. Les régions intrinsèquement désordonnées contiennent fréquemment des motifs courts d’interaction, tels que :
des séquences riches en proline (domaine PRD), comme dans la synapsine impliquée dans l’organisation des vésicules synaptiques,
des répétitions RGG/RG, comme dans la nucléoline impliquée dans l’organisation du nucléole,
des domaines à faible complexité riches en glycine ou en glutamine/asparagine (Q/N), comme dans certaines protéines des granules de stress telles que G3BP1,
des motifs aromatiques (tyrosine, phénylalanine, tryptophane) favorisant des interactions π–π et cation–π avec des résidus chargés, contribuant fortement à la cohésion des condensats, comme dans les domaines intrinsèquement désordonnés des cavines impliquées dans les cavéoles membranaires.
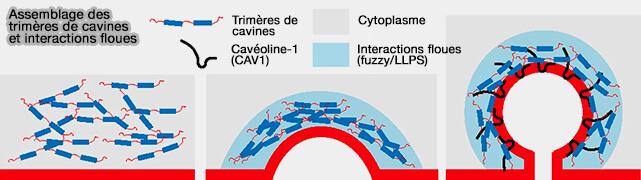
Assemblage des cavines et interactions floues (fuzzi)
(Figure : vetopsy.fr d'après Tillu et coll)
b. Par exemple, les cavines possèdent également des régions intrinsèquement désordonnées, i.e. DR1/23 qui participent à leur oligomérisation et à la génération de courbure membranaire, illustrant l’implication des IDR dans des condensats associés aux membranes ( oligomérisation des cavines).
4. Toutes ces protéines sont aussi impliquées dans les interactions dites " floues ", fuzzy en anglais, qui impliquent des interactions très dynamiques entre des protéines désordonnées ou à ambiguïté structurelle et favorisent des interactions protéine/protéine réversibles et adaptables.
Conséquences : interactions " floues " (fuzzi)
Les interactions dites " floues " (fuzzy en anglais) désignent des modes d’interaction impliquant des protéines intrinsèquement désordonnées ou partiellement structurées, dans lesquels aucune conformation unique et stable n’est imposée.
Ces interactions sont dynamiques, multivalentes et modulables selon le contexte cellulaire, permettant une plasticité structurale compatible avec la formation de réseaux moléculaires adaptatifs.
Ce concept est notamment invoqué pour expliquer l’oligomérisation des cavines et l’organisation dynamique des cavéoles ( oligomérisation des cavines).
1. Les transitions conformationnelles adaptatives associées au flou protéique permettent :
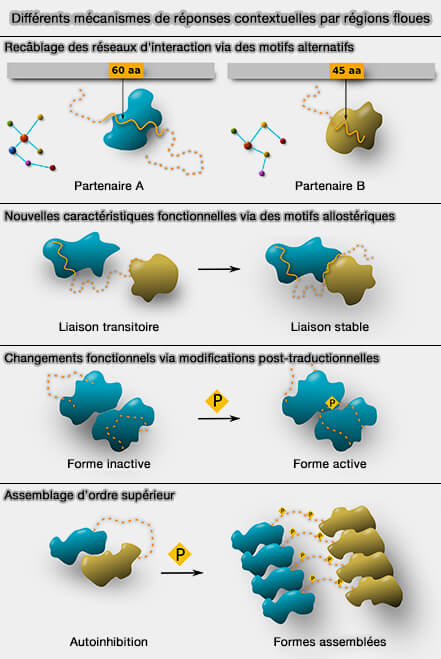
Différentes réponses contextuelles par régions " floues "
(Figure : vetopsy.fr d'après Miskei et coll)
le recâblage des réseaux d'interaction via des motifs alternatifs,
de nouvelles caractéristiques fonctionnelles via des motifs allostériques,
des changements fonctionnels lors de modifications post-traductionnelles,
la régulation de complexes multiprotéiques de signalisation et de contrôle cellulaire.
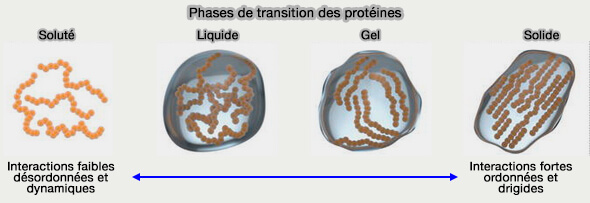
2. La multivalence dynamique permise par ces interactions contribue aux transitions d’état des protéines, autorisant le passage réversible entre états diffus, liquides condensés, viscoélastiques ou, dans certains contextes pathologiques, solides (Protein Phase Separation: A New Phase in Cell Biology 2018).
Sur le plan fonctionnel, cette plasticité moléculaire se traduit par :
la formation de granules de stress (SG), i.e. assemblages de ribonucléoprotéines cytoplasmiques (RNP) à phases séparées, sans membrane, dont la fonction principale est de favoriser la survie des cellules en condensant les ARNm bloqués par la traduction, les composants ribosomiques, les facteurs d'initiation de la traduction et les protéines de liaison à l'ARN (The Integral Role of RNA in Stress Granule Formation and Function 2021),
les corps nucléaires tels que le nucléole, corps de Cajal, les speckles, les paraspecklesou les nuclear stress bodiesn qui représentent des exemples majeurs de compartiments nucléaires organisés par séparation de phase liquide-liquide et interactions multivalentes ARN/protéines,
la détection de changements dans l'homéostasie cellulaire,
Les condensats biomoléculaires associés aux membranes phospholipidiques peuvent générer des contraintes mécaniques locales, favorisant l’induction ou la stabilisation de courbures membranaires ( cavines et courbure membranaire).
2. De nombreuses protéines impliquées dans le trafic membranaire possèdent des domaines intrinsèquement désordonnés volumineux.
Cette augmentation du volume occupé induit un encombrement stérique et des forces entropiques pouvant contribuer à la courbure membranaire ainsi qu’aux processus de fusion ou de fission ( crowding et courbure/fission).







 pathologies des corps nucléaires).
pathologies des corps nucléaires).