


Biologie cellulaire
Mitochondries : génome
Réplication, transcription et maturation des ARN mitochondriaux
- En construction

Les mitochondries possèdent une machinerie d'expression génique spécialisée, héritée de leur origine bactérienne, qui assure la réplication de l'ADNmt, la transcription, la maturation des ARN et la traduction des gènes indispensables à la phosphorylation oxydative.
Bien que reposant sur des principes similaires à ceux de l'expression génique nucléaire, ces mécanismes présentent de nombreuses particularités concernant (Human Mitochondrial RNA Processing and Modifications: Overview 2021).
- la réplication du génome mitochondrial.
- la transcription,
- la maturation des ARN mitochondriaux,
- la traduction par les mitoribosomes.
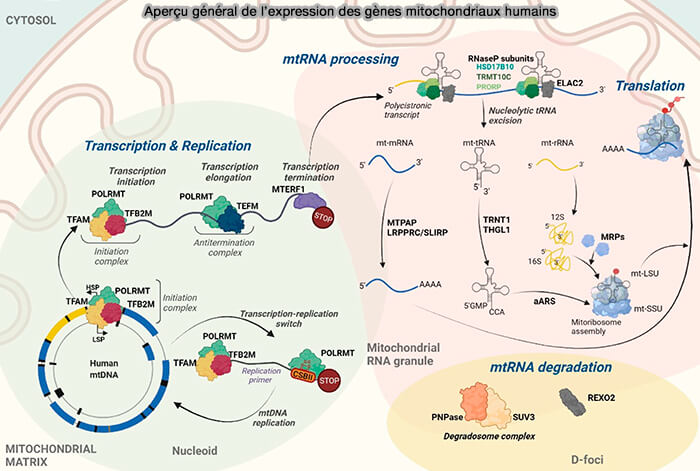
(Figure : vetopsy.fr Jedynak-Slyvka et coll)
Réplication de l'ADN mitochondrial
La réplication de l'ADN mitochondrial permet le maintien du nombre de copies d'ADNmt nécessaires au fonctionnement des mitochondries.

Les mécanismes généraux de la réplication de l'ADN sont longuement détaillés dans des pages spécifiques.
La réplication mitochondriale repose sur les mêmes principes fondamentaux que la réplication de l'ADN nucléaire, mais fait intervenir une machinerie spécifique et un mode de réplication propres aux mitochondries.
1. La réplication de l'ADNmt est assurée par un ensemble de protéines codées par le génome nucléaire, puis importées dans la mitochondrie (Mitochondrial DNA replication in mammalian cells: overview of the pathway 2018).
(Figure : vetopsy.fr Rawi et coll)
a. Cette machinerie comprend principalement :
- l'ARN polymérase mitochondriale (POLRMT), qui synthétise les courtes amorces d'ARN nécessaires à l'initiation de la réplication,
- l'ADN polymérase γ (POL γ), seule ADN polymérase réplicative des mitochondries,
- l'hélicase TWINKLE, qui sépare les deux brins d'ADN (TWINKLE and Other Human Mitochondrial DNA Helicases: Structure, Function and Disease 2020),
- les protéines mtSSB (Mitochondrial Single-Stranded DNA-Binding Protein), qui stabilisent les brins simples d'ADN au cours de la réplication.
b. La réplication débute au niveau de l'origine de réplication du brin lourd (OH, Origin of Heavy-strand replication), où l'ARN polymérase mitochondriale (POLRMT) synthétise une courte amorce d'ARN fournissant l'extrémité 3'-OH nécessaire à l'initiation de la synthèse d'ADN.
L'ADN polymérase γ (POLγ) prend ensuite le relais et synthétise le nouveau brin lourd en utilisant le brin léger comme matrice, tandis que l'hélicase TWINKLE ouvre progressivement la double hélice.
Remarque : contrairement à la réplication de l'ADN nucléaire, où l'amorce d'ARN est synthétisée par le complexe primase-ADN polymérase α, la mitochondrie ne possède ni primase ni ADN polymérase α et cette fonction est assurée par l'ARN polymérase mitochondriale (POLRMT).
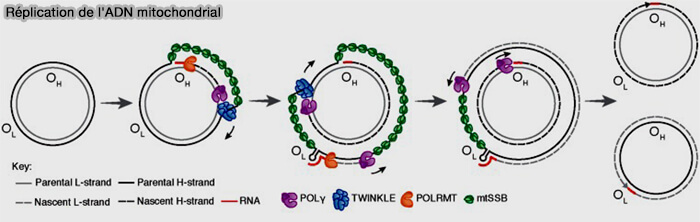
(Figure : vetopsy.fr Rawi et coll)
c. À mesure que la fourche de réplication progresse, l'origine de réplication du brin léger (OL, Origin of Light-strand replication) est exposée.
L'ARN polymérase mitochondriale (POLRMT) y synthétise une nouvelle amorce d'ARN, permettant ensuite à l'ADN polymérase γ (POLγ) d'initier la synthèse du nouveau brin léger en utilisant le brin lourd comme matrice.
d. Après la synthèse des deux nouveaux brins,
- les amorces ARN sont éliminées principalement par la RNase H1 mitochondriale,
- les lacunes sont comblées par l'ADN polymérase γ (POLγ),
- l'ADN ligase III (LIG3), seule ADN ligase présente dans les mitochondries, scelle les dernières discontinuités en formant les dernières liaisons phosphodiester, achevant ainsi la réplication des deux molécules filles d'ADN mitochondrial.
2. La réplication de l'ADNmt est indépendante du cycle cellulaire.
Contrairement à l'ADN nucléaire, l'ADNmt peut être répliqué tout au long du cycle cellulaire afin d'adapter le nombre de copies du génome mitochondrial aux besoins énergétiques de la cellule.
3. Les molécules d'ADNmt sont regroupées dans des complexes nucléoprotéiques appelés nucléoïdes, analogues aux nucléoïdes des procaryotes (Mitochondrial Nucleoid: Shield and Switch of the Mitochondrial Genome 2017).
- Une cellule somatique contient généralement de 100 à 10 000 molécules d'ADNmt réparties entre de nombreux nucléoïdes, chacun renfermant le plus souvent une à huit molécules d'ADN mitochondrial (Functional complementation of mitochondrial DNAs: mobilizing mitochondrial genetics against dysfunction 2010).
- Ces complexes contiennent notamment TFAM, qui joue un rôle essentiel dans la compaction, la protection et l'organisation du génome mitochondrial ainsi que de nombreuses autres protéines (Cross-strand binding of TFAM to a single mtDNA molecule forms the mitochondrial nucleoid 2015).
- Ils constituent également des plateformes où sont coordonnées la réplication, la transcription et le maintien de l'intégrité de l'ADNmt (Mitochondrial Nucleoid: Shield and Switch of the Mitochondrial Genome 2017).
Remarque : les plastes des végétaux, notamment les chloroplastes, possèdent également des nucléoïdes, qui organisent et compactent leur ADN.
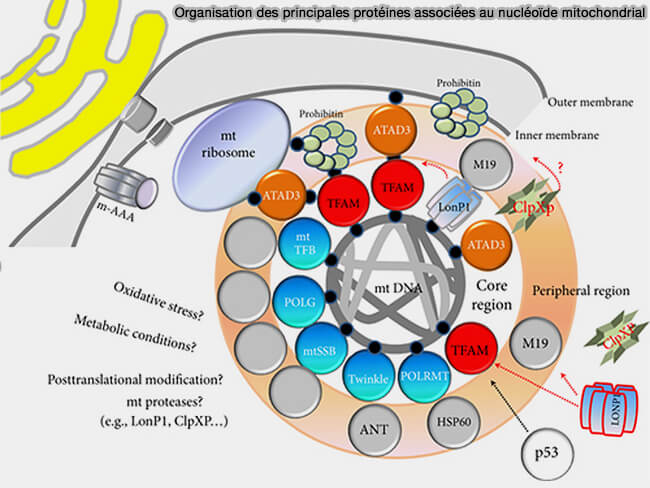
(Figure : vetopsy.fr Lee et Han)
4. La réplication indépendante des nombreuses copies d'ADNmt favorise l'apparition, l'accumulation et la transmission de mutations au sein d'une même cellule.
- On appelle hétéroplasmie la coexistence, dans une même cellule, d'ADNmt normal et d'ADNmt muté.
- La proportion relative des différentes molécules d'ADNmt peut varier au cours des divisions cellulaires et influence directement la sévérité de nombreuses maladies mitochondriales.
Transcription mitochondriale
La transcription mitochondriale repose sur une machinerie minimale, codée par le génome nucléaire puis importée dans la matrice, qui lit l'ADNmt à partir de promoteurs situés dans la région de contrôle.
- Contrairement à la transcription nucléaire, elle ne produit pas des ARN correspondant à un seul gène, mais de longs transcrits polycistroniques renfermant successivement les séquences des futurs ARNm, ARNt et ARNr mitochondriaux.
- Ces précurseurs seront ensuite clivés et maturés afin de libérer les ARN fonctionnels.
Les mécanismes généraux de la transcription sont longuement détaillés dans des pages spécifiques.
La transcription mitochondriale repose sur les mêmes principes fondamentaux :
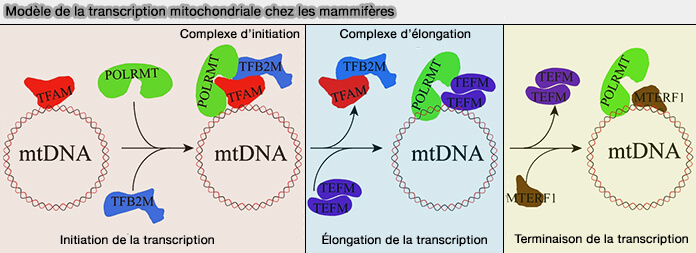
(Figure : vetopsy.fr Lei et coll)
Cependant elle met en œuvre une machinerie distincte et présente plusieurs particularités liées à l'organisation et à l'origine bactérienne du génome mitochondrial.
Initiation de la transcription
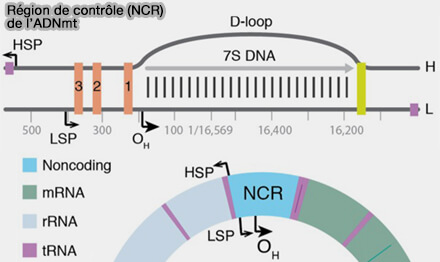
1. Les promoteurs mitochondriaux sont situés dans la région de contrôle ou D-loop, principale région non codante de l'ADNmt (Structure, mechanism, and regulation of mitochondrial DNA transcription initiation 2020).
a. Le promoteur du brin léger, LSP (Light Strand Promoter), permet la transcription du brin léger de l'ADN mitochondrial, qui contient notamment le gène ND6 ainsi que plusieurs gènes d'ARNt.
Le transcrit initié à partir de ce promoteur présente une particularité, car il peut également fournir l'amorce ARN nécessaire à l'initiation de la réplication du brin lourd.
b. Les promoteurs du brin lourd comprennent HSP1 et HSP2 (Heavy Strand Promoters).
- HSP1 assure principalement la transcription des gènes codant les ARNr 12S et 16S ainsi que des ARNt adjacents, nécessaires à la biogenèse des mitoribosomes.
- HSP2 permet la transcription polycistronique d'une grande partie du brin lourd, à l'origine des ARNm codant la majorité des protéines mitochondriales de la chaîne respiratoire, ainsi que de plusieurs ARNt.

Vous pouvez lire : Structural basis of mitochondrial transcription (2019) et Structural basis of mitochondrial transcription initiation (2019).
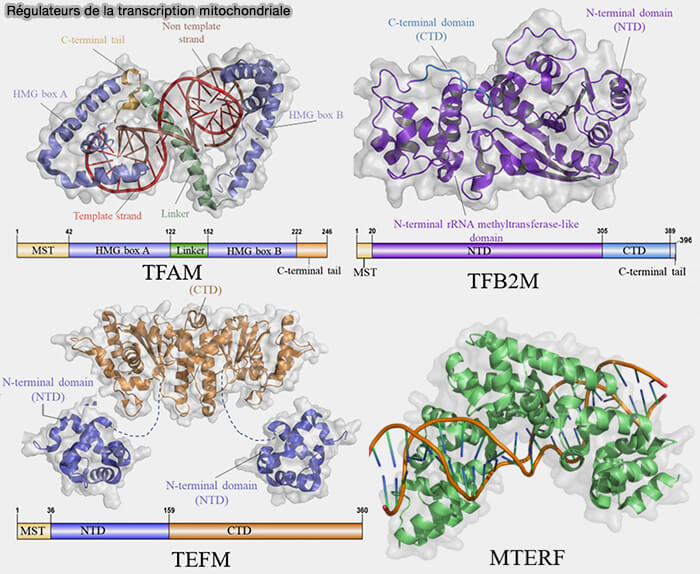
2. TFAM (Mitochondrial Transcription Factor A) reconnaît et se fixe en amont des trois promoteurs mitochondriaux, LSP, HSP1 et HSP2.
- Selon le promoteur recruté, il initie la transcription du brin léger (LSP) ou du brin lourd (HSP1 et HSP2).
- Il induit une courbure marquée de l'ADNmt qui facilite le recrutement de POLRMT et favorise l'assemblage du complexe d'initiation transcriptionnelle.
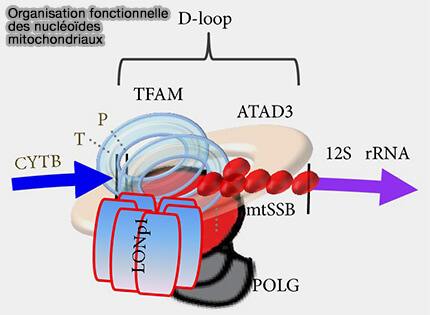
(Figure : vetopsy.fr Lee et Han)
a. Selon son abondance et son mode de fixation, TFAM exerce cependant une seconde fonction architecturale en compactant l'ADN mitochondrial.
- Grâce à ses deux domaines HMG-box (A et B), elle se fixe dans le sillon mineur de la double hélice et induit une courbure très marquée de l'ADN, pouvant atteindre près de 180°, qui replie localement la molécule en une structure en fer à cheval ou en U.
- Chaque molécule de TFAM recouvre ainsi environ 15 à 20 paires de bases de l'ADNmt.
- La fixation successive de centaines de molécules de TFAM provoque la formation de multiples courbures et boucles qui se stabilisent par les interactions entre ces protéines, conduisant à la compaction tridimensionnelle de l'ADNmt au sein des nucléoïdes mitochondriaux.
b. Cette organisation reste dynamique.
- La compaction correspond à l'état P (Packaging) représenté sur la figure, dans lequel l'ADNmt est principalement condensé et moins accessible à la transcription et protège le génome mitochondrial.
- À l'inverse, l'état T (Transcription) correspond à une configuration plus ouverte des promoteurs, permettant le recrutement de POLRMT, l'assemblage du complexe d'initiation et le démarrage de la transcription.
c. La figure illustre également plusieurs protéines associées aux nucléoïdes mitochondriaux.
- LONP1 dégrade TFAM lorsqu'il n'est pas lié à l'ADNmt, contribuant ainsi à l'homéostasie des nucléoïdes.
- POLRMT est recrutée au niveau des promoteurs pour assurer la transcription.
- POLγ, TWINKLE et mtSSB interviennent lors de la réplication de l'ADN mitochondrial.
- ATAD3 est une protéine de la membrane interne mitochondriale qui contribue à l'organisation et à l'ancrage des nucléoïdes à proximité de la membrane interne.
3. La transcription de l'ADN mitochondrial est réalisée par une unique ARN polymérase mitochondriale, POLRMT, une enzyme constituée d'une seule sous-unité, apparentée aux ARN polymérases des bactériophages T3 et T7, mais distincte des ARN polymérases nucléaires Pol I, Pol II et Pol III (Mitochondria transcription and cancer 2024).
a. POLRMT contient :
- une extension N-terminale (NTE), un domaine PPR (Pentatricopeptide Repeat) et un domaine N-terminal (NTD), régions qui participent principalement aux interactions avec les facteurs de transcription mitochondriaux, à la reconnaissance des promoteurs et à la formation du complexe d'initiation transcriptionnelle.
- un domaine C-terminal (CTD) qui constitue le domaine catalytique de l'enzyme et renferme le site actif responsable de la polymérisation des ribonucléotides au cours de la synthèse de l'ARN.
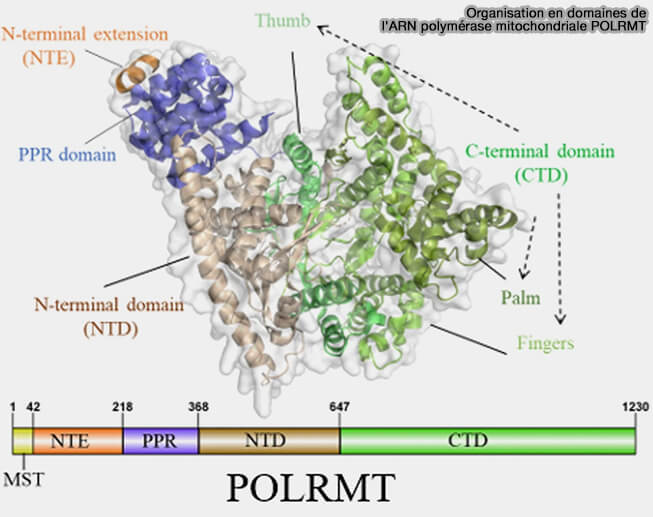
(Figure : vetopsy.fr Lei et coll)
b. POLRMT est codée par le génome nucléaire puis importée dans la matrice mitochondriale.
POLRMT est recrutée au niveau des promoteurs mitochondriaux par TFAM, mais elle ne peut initier seule la transcription.
4. TFB2M (Mitochondrial Transcription Factor B2) intervient après le recrutement de POLRMT en favorisant l'ouverture locale du duplex d'ADN au niveau du promoteur et en stabilisant le complexe d'initiation transcriptionnelle (Human mitochondrial transcription factors TFAM and TFB2M work synergistically in promoter melting during transcription initiation 2016).
Cette étape permet l'engagement de POLRMT sur le brin matrice et le démarrage de la synthèse de l'ARN mitochondrial à partir de l'un des trois promoteurs mitochondriaux (LSP, HSP1 ou HSP2).
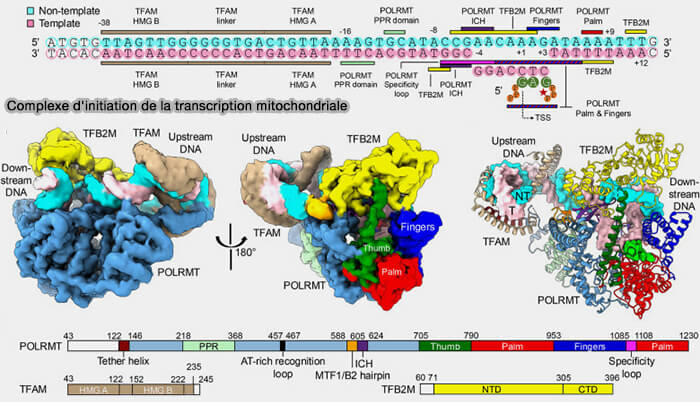
(Figure : vetopsy.fr Shen et coll)
Élongation de la transcription
Après son initiation au niveau de l'un des promoteurs mitochondriaux (LSP, HSP1 ou HSP2), POLRMT entre en phase d'élongation ( principes généraux de l'élongation).
principes généraux de l'élongation).
Vous pouvez lire : Mechanism of transcription anti-termination in human mitochondria (2018).
(Figure : vetopsy.fr Lei et coll)
Élongation des transcrits mitochondriaux
1. Cette transition vers l'élongation est facilitée par TEFM (Transcription Elongation Factor of Mitochondria), qui se fixe sur POLRMT et augmente sa processivité en limitant les arrêts prématurés de la transcription (TEFM is a potent stimulator of mitochondrial transcription elongation in vitro 2015).
2. Après son initiation, POLRMT progresse le long du brin matrice correspondant et synthétise un ARN dans le sens 5' ➞ 3' (TEFM is a potent stimulator of mitochondrial transcription elongation in vitro 2015).
Selon le promoteur utilisé, la transcription concerne le brin lourd (HSP1 et HSP2) ou le brin léger (LSP).
Couplage entre transcription et réplication au niveau de LSP
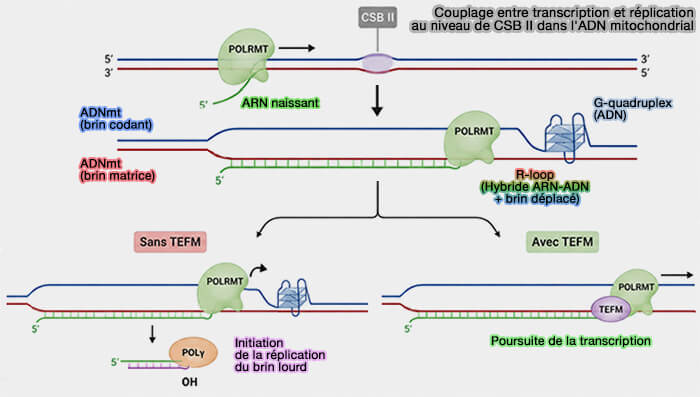
Lorsque la transcription est initiée à partir du promoteur LSP, elle présente une particularité importante qui assure le couplage entre transcription et réplication de l'ADN mitochondrial.
1. En présence de TEFM, POLRMT traverse la région CSB II (Conserved Sequence Block II) située dans la région de contrôle et poursuit la transcription sur une grande partie du génome mitochondrial, produisant de longs ARN polycistroniques (TEFM facilitates transition from RNA synthesis to DNA synthesis at H-strand replication origin of mtDNA 2025).
(Figure : vetopsy.fr)
2. En revanche, en l'absence de TEFM, POLRMT a tendance à s'arrêter au niveau de CSB II.
a. CSB II correspond à une région de l'ADN mitochondrial dont le brin codant est particulièrement riche en guanines.
Lors de la transcription initiée à partir du promoteur LSP, POLRMT lit le brin matrice complémentaire, riche en cytosines, et synthétise ainsi un ARN lui-même riche en guanines.
- Cet ARN, au lieu d'être rapidement libéré, reste apparié au brin matrice d'ADN, empêchant la réassociation normale des deux brins d'ADN derrière la polymérase et il se forme ainsi un hybride ARN-ADN stable appelé R-loop.
- Le second brin d'ADN, déplacé et maintenu sous forme simple brin, se replie alors en une structure secondaire stable appelée G-quadruplex, résultant de l'association de quatre guanines (G-quadruplexes and their regulatory roles in biology 2015).
Ces deux structures constituent un obstacle à la progression de POLRMT et favorisent l'arrêt prématuré de la transcription.
b. Le court ARN ainsi produit reste associé à l'ADN mitochondrial et sert d'amorce à l'ADN polymérase γ (POLγ) pour initier la réplication du brin lourd à partir de l'origine OH.
3. Ainsi, la transcription initiée à partir du promoteur LSP peut être orientée :
- soit vers la production complète des ARN mitochondriaux,
- soit vers la synthèse des amorces ARN nécessaires à la réplication, établissant un lien étroit entre transcription et réplication de l'ADN mitochondrial.
4. Les longs transcrits obtenus contiennent successivement les séquences des futurs ARNm, ARNt et ARNr mitochondriaux.
Ils sont ensuite pris en charge par les mécanismes de maturation, qui assurent leur clivage et leur maturation pour produire les ARNm, ARNt et ARNr fonctionnels.
Terminaison de la transcription
La terminaison de la transcription mitochondriale correspond à l'arrêt de la synthèse de l'ARN par POLRMT et à la libération du transcrit nouvellement synthétisé ( principes généraux de la terminaison).
Contrairement à l'initiation de la transcription, ses mécanismes restent incomplètement élucidés et semblent varier selon les promoteurs et les transcrits concernés.
1. Plusieurs protéines appartenant à la famille des MTERF (Mitochondrial Transcription Termination Factors) participent à la régulation de l'expression du génome mitochondrial.
a. Chez les mammifères, cette famille comprend quatre protéines homologues, MTERF1, MTERF2, MTERF3 et MTERF4, caractérisées par la présence de répétitions structurales mTERF leur permettant de reconnaître des séquences spécifiques de l'ADN.
b. Malgré leur dénomination, ces protéines ne sont pas toutes impliquées dans la terminaison de la transcription :
- MTERF1 intervient principalement dans la terminaison de la transcription et contribue à limiter les interférences entre transcription et réplication.
- MTERF2 participe à la régulation de l'expression des gènes mitochondriaux, influençant ainsi l'assemblage et l'activité des complexes de la chaîne respiratoire.
- MTERF3 agit comme un régulateur négatif de l'initiation de la transcription.
- MTERF4 forme avec la méthyltransférase NSUN4 un complexe impliqué dans la maturation des ARNr mitochondriaux et le contrôle de l'assemblage de la grande sous-unité du mitoribosome.
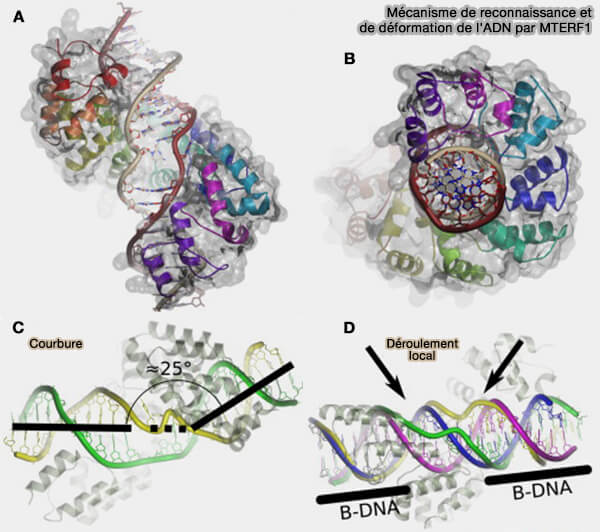
2. MTERF1, facteur de terminaison le mieux caractérisé, se fixe avec une forte affinité sur une séquence spécifique localisée dans le gène MT-TL1, codant l'ARNtLeu (UUR), immédiatement en aval du gène de l'ARNr 16S (Helix Unwinding and Base Flipping Enable Human MTERF1 to Terminate Mitochondrial Transcription 2010).
(Figure : vetopsy.fr Yakubovskaya et coll)
a. La liaison de MTERF1 à l'ADN (A et B) induit une courbure de la double hélice (C), accompagnée d'un déroulement local du duplex au niveau du site de liaison (D).
Cette déformation du duplex constitue une barrière physique à la progression de POLRMT, provoquant l'arrêt de la transcription et la libération du transcrit.
b. Ce mécanisme contribue à délimiter l'unité transcriptionnelle contenant les deux ARNr mitochondriaux et participe à la régulation de leur expression.
Remarque : au-delà de son rôle dans la terminaison de la transcription, MTERF1 pourrait également coordonner la progression des complexes de transcription et de réplication en agissant comme une barrière polaire limitant les collisions entre les deux machineries (Mitochondrial transcription termination factor 1 directs polar replication fork pausing 2016).
3. La terminaison des longs transcrits polycistroniques issus des promoteurs HSP2 et LSP est moins bien comprise et résulte probablement de l'action combinée de séquences d'ADN, car aucune séquence consensus de terminaison comparable aux terminateurs bactériens n'a été clairement identifiée, de structures formées par les ARN naissants et de facteurs régulateurs spécialisés.
Les transcrits polycistroniques obtenus ne sont pas directement fonctionnels et doivent être :
- clivés selon le modèle de ponctuation par les ARNt,
- maturés afin de générer les ARNm, ARNt et ARNr mitochondriaux fonctionnels.
Maturation des ARN mitochondriaux
La maturation des ARN mitochondriaux transforme les longs transcrits polycistroniques produits par POLRMT en ARN fonctionnels et repose sur :
- des clivages précis,
- des modifications post-transcriptionnelles,
- des étapes de stabilisation propres aux ARNm, ARNt et ARNr mitochondriaux.
Les transcrits mitochondriaux primaires contiennent des séquences d'ARNm, d'ARNt et d'ARNr disposées les unes à la suite des autres selon l'organisation compacte de l'ADNmt.
Clivage et maturation des ARNt mitochondriaux (ARNtmt)
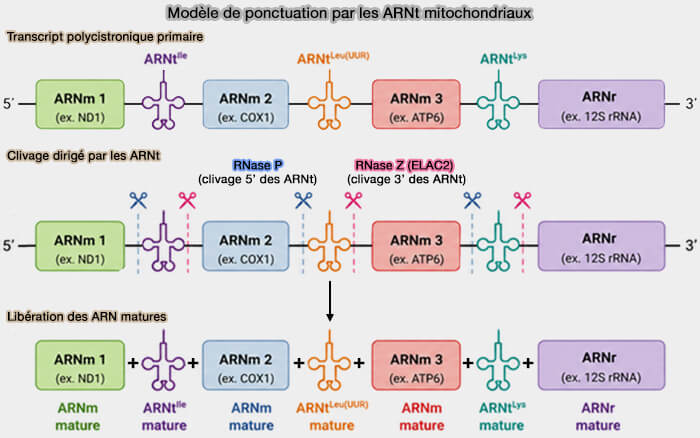
Le modèle de ponctuation par les ARNt (tRNA punctuation model), proposé en 1981, établit que les ARNt intercalés entre la plupart des gènes mitochondriaux servent de repères moléculaires pour le clivage des transcrits polycistroniques (tRNA punctuation model of RNA processing in human mitochondria 1981)
Ce modèle explique l'essentiel de la maturation des ARN mitochondriaux, bien que plusieurs exceptions et mécanismes complémentaires aient été identifiés par la suite.
(Figure : vetopsy.fr)
2. Après leur excision des transcrits polycistroniques, les pré-ARNt mitochondriaux subissent une succession d'étapes de maturation coordonnées impliquant plusieurs enzymes spécialisées (Structural basis for human mitochondrial tRNA maturation 2024)
a. Le complexe TRMT10C-SDR5C1 reconnaît le pré-ARNt, contribue à sa stabilisation et catalyse la méthylation de certains nucléotides, facilitant ainsi le positionnement du substrat pour les enzymes intervenant aux étapes suivantes (Structural basis of RNA processing by human mitochondrial RNase P 2021).
b. Le clivage en 5' des pré-ARNt mitochondriaux est assuré par la RNase P mitochondriale, complexe protéique qui, outre TRMT10C (MRPP1) et SDR5C1 (MRPP2), contient PRORP (MRPP3) qui porte l'activité catalytique de clivage.
Contrairement à la RNase P nucléaire, elle est dépourvue d'ARN catalytique et est constituée exclusivement de protéines.
c. Le clivage en 3' des pré-ARNt mitochondriaux est ensuite réalisé par ELAC2, homologue mitochondrial de la RNase Z.
- Comme les autres RNases Z eucaryotes, ELAC2 reconnaît l'extrémité 3' des pré-ARNt et élimine les séquences surnuméraires situées en aval de l'ARNt mature.
- Ce clivage prépare l'ajout de la séquence terminale CCA.
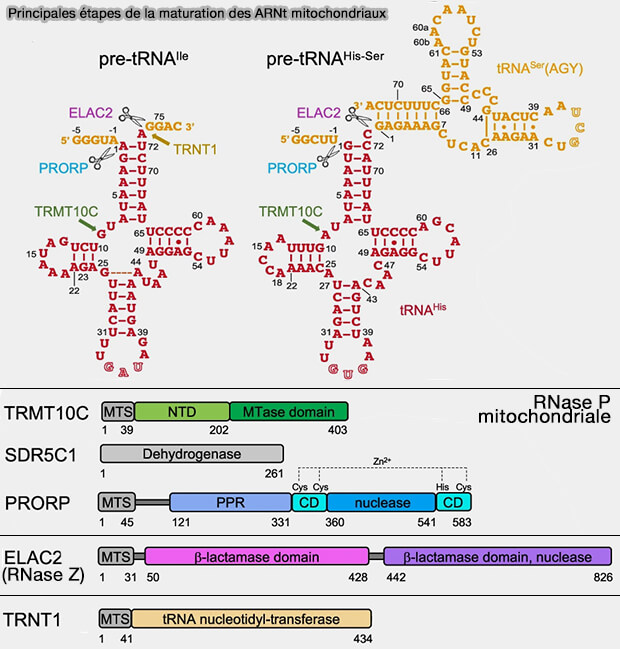
(Figure : vetopsy.fr d'après Meynier et coll)
d. Enfin, TRNT1 ajoute, sans matrice, la séquence terminale CCA à l'extrémité 3', indispensable à l'aminoacylation des ARNtmt, qui permet la fixation de l'acide aminé correspondant sur l'adénosine terminale, condition nécessaire à la traduction mitochondriale.
Ces différentes étapes libèrent progressivement les ARNt mitochondriaux et séparent simultanément les ARNm et les ARNr voisins.
3. Les ARNt mitochondriaux subissent ensuite de nombreuses modifications post-transcritionnelles portant sur leurs bases ou leur ribose :
- l'isomérisation de l'uridine en pseudouridine (Ψ),
- la réduction de l'uridine en dihydrouridine (D),
- la désamination de l'adénosine en inosine (I),
- diverses méthylations de bases, telles que la 1-méthyladénosine (m1A), la 5-méthylcytidine (m5C) ou la 7-méthylguanosine (m7G ),
- la 2′-O-méthylation du ribose (Nm).
4. Certaines modifications, affectant notamment la position wobble de l'anticodon, sont particulièrement importantes dans les ARNtmt, car elles contribuent à la reconnaissance correcte des codons mitochondriaux et compensent les particularités structurales de plusieurs ARNt mitochondriaux (Human Mitochondrial RNA Processing and Modifications: Overview 2021).
En effet, plusieurs ARNt mitochondriaux présentent des structures atypiques minimalistes par rapport aux ARNt cytosoliques.
Certains ont notamment perdu l'une des boucles caractéristiques de la structure en trèfle classique, comme l'ARNtSer(AGY) dépourvu de boucle D, et dépendent davantage des protéines de maturation et de traduction mitochondriales pour assurer leur stabilité et leur fonctionnement ( figure au-dessus).
Maturation des ARNm et des ARNr mitochondriaux
1. Les ARNm mitochondriaux, libérés des transcrits polycistroniques par le modèle de ponctuation des ARNt :
- ne possèdent généralement pas de coiffe 5',
- ne subissent pas d'épissage classique.
a. Ils sont stabilisés par des protéines de liaison aux ARN mitochondriaux.
b. Ils acquièrent souvent une queue poly(A), ajoutée par la poly(A) polymérase mitochondriale MTPAP.
- Contrairement aux ARNm nucléaires, cette polyadénylation ne dépend pas du complexe de clivage et de polyadénylation (CPA), ni de la poly(A) polymérase nucléaire PAP, mais d'une machinerie mitochondriale spécifique.
- La polyadénylation mitochondriale stabilise certains ARNm et peut aussi compléter des codons stop incomplets présents à l'extrémité 3' de plusieurs transcrits, l'ajout de résidus adénosine transformant alors un codon U ou UA terminal en codon stop UAA fonctionnel.
2. Les ARNr mitochondriaux 12S et 16S sont également libérés à partir des transcrits primaires, puis modifiés et assemblés avec des protéines mitoribosomiques codées par le génome nucléaire.
- Ces ARNr ne constituent qu'une fraction limitée de la masse du mitoribosome, beaucoup plus riche en protéines que le ribosome cytosolique.
- Leur maturation conditionne l'assemblage correct des mitoribosomes et donc la synthèse des protéines codées par l'ADN mitochondrial.

La maturation des ARN mitochondriaux est donc étroitement couplée à la traduction mitochondriale et tout défaut de clivage, de modification ou de stabilisation de ces ARN peut compromettre la synthèse des protéines codées par l'ADNmt et perturber l'assemblage des complexes de la phosphorylation oxydative.
Traduction mitochondriale et assemblage
des complexes respiratoires
Les ARNm, ARNt et ARNr mitochondriaux matures sont ensuite utilisés par les mitoribosomes pour assurer la traduction des protéines codées par l'ADN mitochondrial, leur insertion dans la membrane interne et l'assemblage des complexes de la chaîne respiratoire.
Biologie cellulaire et moléculaireMembrane plasmiqueNoyauCytoplasmeRéticulum endoplasmiqueAppareil de GolgiMitochondriesEndosomesLysosomesPeroxysomesProtéasomesCytosqueletteFilaments d'actineFilaments intermédiairesMicrotubulesMatrice extracellulaireReproduction cellulaireBiochimieTransport membranaire Moteurs moléculairesVoies de signalisation