les RIM (Regulating synaptic membrane exocytosis protein), en particulier RIM1α, formerait une attache longue, raccourcie par des interactions avec d'autres protéines, i.e. les RIM-BP (RIM-Binding Protein), ELKS/CAST, la liprine-α,
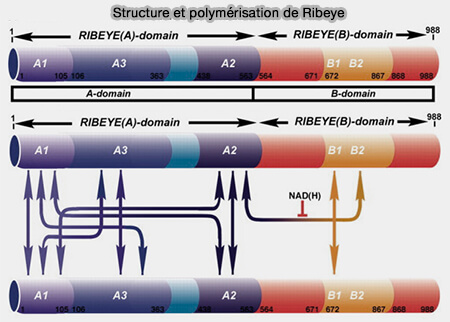
Structure et polymérisation de Ribeye
(Figure : vetopsy.fr d'après Magupalli et coll)
1. Ribeye est constitué d'un domaine A spécifique N-terminal et d'un domaine B C-terminal qui est identique au CtBP2 du corepresseur transcriptionnel, à l'exception des 20 premiers acides aminés.
Le domaine B de Ribeye se lie à NAD+/NADH avec une affinité élevée.
2. L'expression de RIBEYE in vitro est suffisante pour la formation d'agrégats sphériques non structurés, car RIBEYE peut s'auto-assembler, via cinq sites distincts, par des interactions hétérotypiques entre ses domaines A et B.
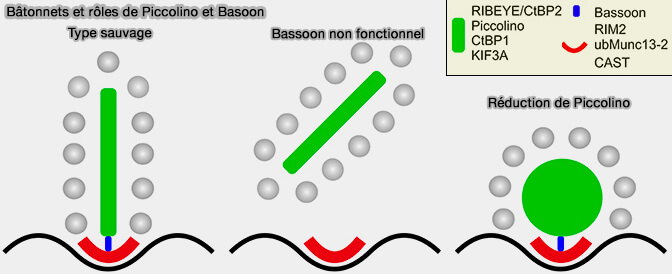
Délétion de Ribeye dans les synapses à ruban
(Figure : vetopsy.fr d'après Maxeiner et coll)
Par exemple, l'interaction de PIccolino avec Ribeye joue un rôle important dans l'assemblage et l'ancrage des rubans synaptiques (
3. Cependant, la seule expression de RIBEYE ne suffit pas à créer les SR allongés en forme de plaque comme, par exemple, dans les photorécepteurs vertébrés ou les cellules bipolaires de la tige de la rétine.
Dans la rétine, le rapport Ribeye/Piccolino est de 1:1.
Dans l'oreille, le nombre de molécules de Ribeye est bien supérieur à celui de Piccolino, pouvant empêcher Piccolino d'imposer une disposition régulière des protéines Ribeye en forme de plaque .
Bâtonnets et rôles de Piccolino et Basoon
(Figure : vetopsy.fr d'après Regus-Leidig et coll)
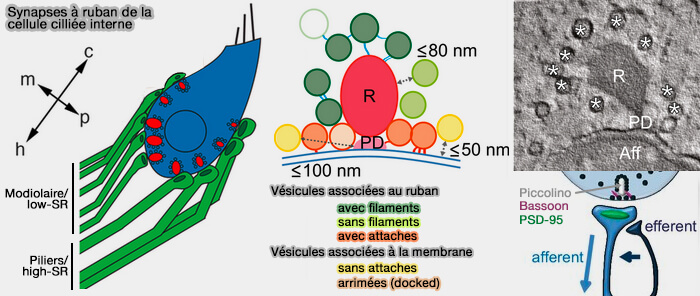
L'amarrage (docking) et l'amorçage (priming) qui préparent les vésicules synaptiques pour la fusion avec la membrane présynaptique et la libération du neurotransmetteur lors d'un afflux calcique.
1. Une séquence analogue à celle des synapses classiques semble se dérouler également dans les synapses à ruban, en particulier avec le complexe SNARE.
a. Contrairement aux synapses neuronales centrales conventionnelles, les cellules ciliées internes matures (CCI) sont dépourvues des senseurs (ou capteurs) calciques classiques, i.e. synaptotagmine I/II, i.e. senseur calcique, et Munc13, qui jouent un rôle crucial dans l'exocytose des vésicules.
L'oterfiline les remplace dans les IHC, en jouant un rôle primordial et en se liant :
à la membrane vésiculaire,
aux canaux calciques (Cav1.3),
aux protéines SNARE sur la membrane présynaptique
2. Toutefois, les taux de reconstitution des VS sont extrêmement rapides et il est difficile d'analyser ces phénomènes.



{kind=link}


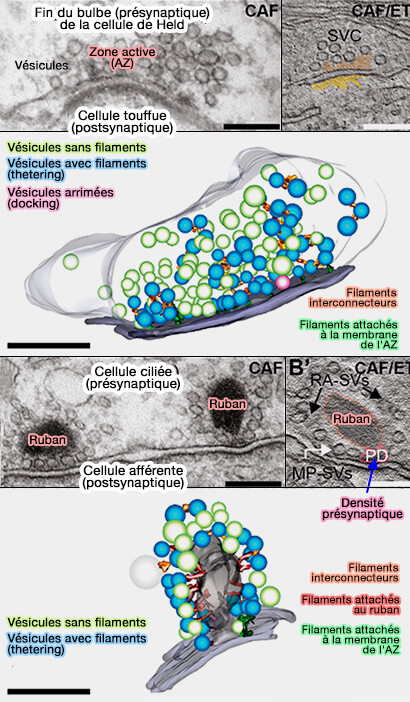
 1. translocation des VS).
1. translocation des VS).