Trafic vésiculaire
Voie sécrétoire
Modèles du trafic intra-Golgien (IGT) : DM et KAR
Sommaire
De nombreux modèles différents et leurs combinaisons ont été proposés pour expliquer les données expérimentales sur le transport réticulum/Golgi (EGT).
À l'heure actuelle, quatre principaux modèles de transport intracellulaire reposant sur trois grands principes, i.e. dissociation, progression et diffusion, sont décrits :
Cette libération conduit à la fusion de vésicules COPI enrichies en Qb SNARE avec des citernes de Golgi, et à la restauration du complexe SNARE du Golgi.
2. Toutefois, des observations contredisent le DM :
la concentration des cargos diffusibles, i.e. les gradients protéiques, lipidiques et ioniques à travers les empilements de saccules du Golgi,
l'importance des protéines SNARE à chaque étape du transport, sans justification car il n'y a ni fusion, ni fission,
la rareté des connexions,
la présence d'empilements de saccules sans connections pendant le transport,
1. La concentration des cargos et mégacargos est supérieure du côté trans du Golgi.
2. Les différents cargos et mégacargos membranaires forment des domaines différents.
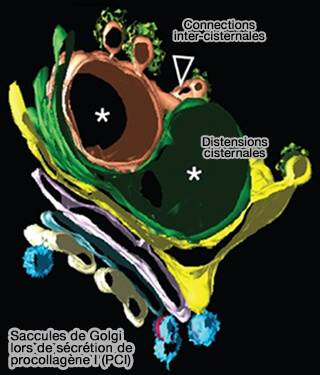
Les distensions des saccules contenant des mégacargos représentent déjà des domaines de fret.
Des pores peuvent être observés entre les distensions contenant le PCI et le reste des citernes.
Les domaines de fret sont séparés du reste des citernes de Golgi par des rangées de pores ou de tubules minces, comme sites de fission ultérieure, même si les fonctions des pores restent inconnues.
3. Les protéines SNARE sont concentrées dans ces domaines de fret, i.e. pendant l'IGT, les SNARE travaillant du côté cis du Golgi enrichies sur les domaines de fret sont remplacées par des SNARE qui fonctionnent du côté trans du complexe de Golgi.
La concentration des cargos et des SNARE augmente l'efficacité.
Les pores ou les tubules minces derrière un domaine de cargaison fournissent la direction.
Golgi lors de sécrétion de procollagène I (PCI)
(Figure : vetopsy.fr d'après Mironov et Beznoussenko)
GS27 et GS28 sont enrichies sur les vésicules COPI du Golgi, alors que Bet1, GS15, Ykt6 et la syntaxine 5 (Stx5) étaient appauvries. Ykt6, Bet1 et GS15 sont enrichis sur des domaines de fret, Stx5 sur les bords des saccules conventionnels.
Au cours de l'IGT, la concentration de cis-SNARE sur les domaines de fret a augmenté.
Puis, les cis-SNARE ont été remplacés par les trans-SNARE.
Les SNARE ont été enrichis sur les domaines de fret situés dans la citerne de Golgi proximale la plus proche.
La plupart des distensions des citernes de Golgi contenant des cargos sont séparées du reste des citernes de Golgi par des pores.
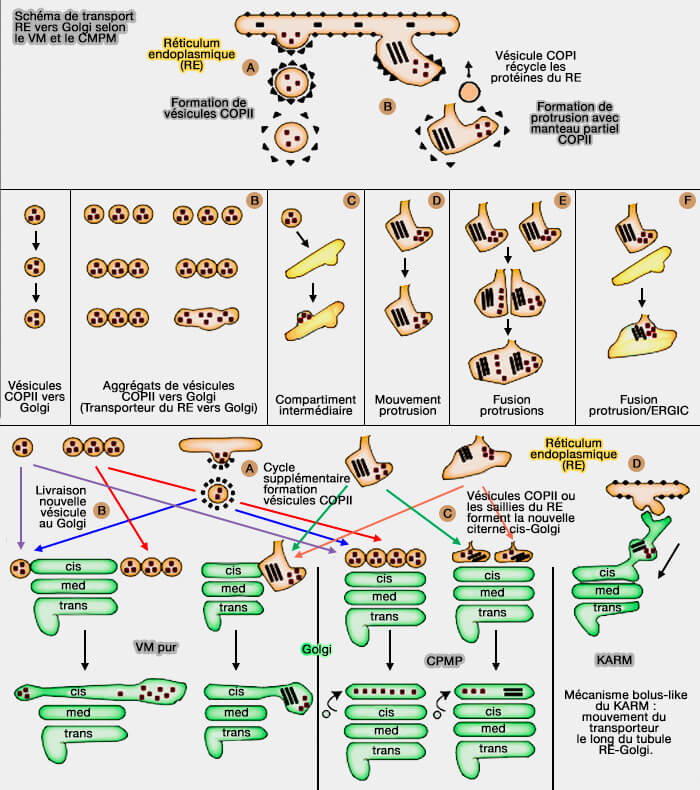
4. Le nombre de citernes du Golgi médian est constant, indépendamment de la quantité de cargos se déplaçant à travers le complexe de Golgi.
Le CMPM qui présuppose qu'après l'arrivée soudaine de différentes quantités de cargaison, le nombre de citernes de Golgi médianes peut augmenter en fonction de ces quantités de cargos.
L'arrivée des cargos augmente la longueur, la surface et le volume des saccules, bien que le nombre de citernes médiales soit resté constant.
5. Les pores cisternaux sont consommés pendant l'IGT.
Après leur formation COPI-dépendante et leur fusion consécutive avec les citernes, les vésicules COPI génèrent l'excès de courbure membranaire qui conduit à la formation de tubules puis à la fusion arrière de la pointe avec la citerne, ce qui pourrait restaurer les pores cisternaux.
COPI participe également à la fission des tubules qui séparent les pores dans la rangée de pores.
7. L'observation principale en contradiction avec le KARM provient du fait que les compartiments séparés du Golgi ne seraient pas attachés les uns aux autres et ne pourraient donc pas fusionner.
En effet, chez S. cerevisiae, tous les compartiments de Golgi sont localisés loin les uns des autres.
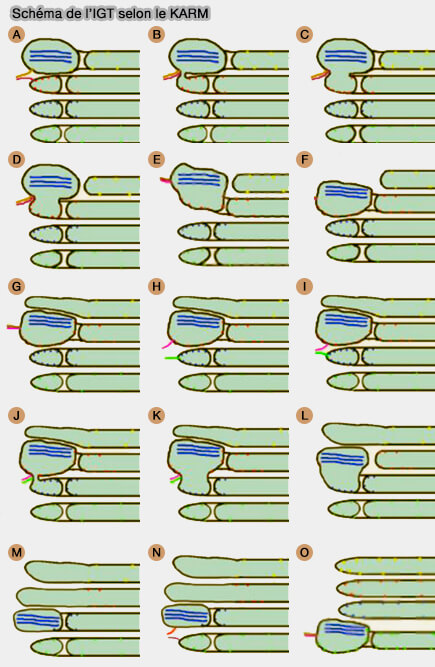
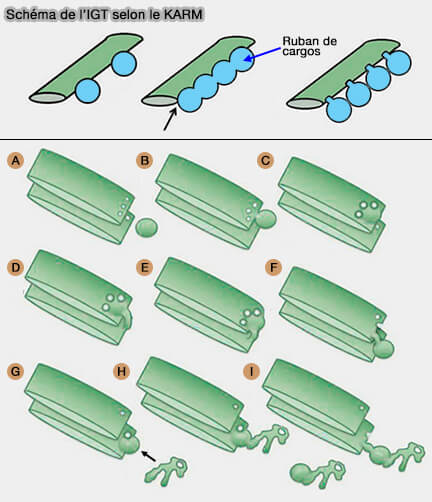
Le mécanisme de l'IGT selon le KARM serait le suivant.
1. La formation du ruban (ribbon en anglais) de cargos s'effectue par :
l'arrivée des domaines cargo et de leur fusion conduisant à un ruban parallèle,
le bourgeonnement des domaines de fret des citernes de Golgi.
2. Dans le KARM asymétrique, le domaine cargo fusionne avec les rebords cisternaux contenant plusieurs pores (B-C).
Le domaine cargo fusionne avec la citerne distale avec le tubule (DE)
Les tubules dans la zone où se trouvaient les pores se disloquent (F).
Le tubule reliant le domaine cargo et la citerne proximale subit une fission (G).
Puis, on assiste à l'arrivée du TGN, sa fusion avec le domaine cargo (H) et la fission consécutive du tubule reliant le domaine cargo et la citerne distale (I).






 principes de base du KARM).
principes de base du KARM).