


Trafic vésiculaire
Voie sécrétoire
Modèles généraux
- En construction

Le transport intracellulaire est l'un des problèmes les plus déroutants dans le domaine de la biologie cellulaire. De nombreux modèles différents et leurs combinaisons ont été proposés pour expliquer les données expérimentales sur le transport intracellulaire.
Ce chapitre se concentre sur les mécanismes supposés du transport du réticulum endoplasmique (RE) vers l'appareil de Golgi et intra-Golgien. On parle :

Les trafics endocytaires et endo-lysosomal sont traités dans des chapitres spéciaux.
À l'heure actuelle, quatre principaux modèles de transport intracellulaire reposant sur trois grands principes, i.e. dissociation, progression et diffusion, sont décrits :
- le modèle vésiculaire (VM),
- le modèle de maturation-progression des citernes (CMPM),
- le modèle de diffusion (DM),
- le modèle kiss-and-run (KARM), qui existe sous forme de deux variantes, symétrique et asymétrique.

Vous pouvez lire l'article qui a inspiré ce chapitre : Models of Intracellular Transport: Pros and Cons (2019).
Vue d'ensemble des modèles
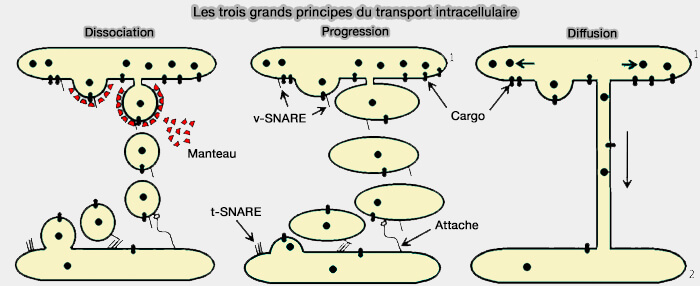
Les modèles reposent sur trois grands principes du transport intracellulaire.
1. La dissociation, principalement vésiculaire, repose sur la formation initiale de bourgeons membranaires dotés d'une enveloppe protéique sur un compartiment proximal.
- Après la fission et le détachement, les vésicules dépendantes de l'enveloppe se déplacent vers un compartiment distal où elles sont amarrées par un système d'attache.
- Grâce aux protéines SNARE, la vésicule fusionne avec le deuxième compartiment.
(Figure : vetopsy.fr d'après Mironov et Beznoussenko)
2. Le mode de progression est fondé sur la formation d'une grande saillie membranaire formée à partir d'un compartiment proximal qui subit une fission.
- Ce grand transporteur est capturé par le système d'attache.
- Grâce aux protéines SNARE, il subit une fusion avec un compartiment distal.
3. Le mode de diffusion du transport intracellulaire fait intervenir une communication entre un compartiment proximal et un compartiment distal.
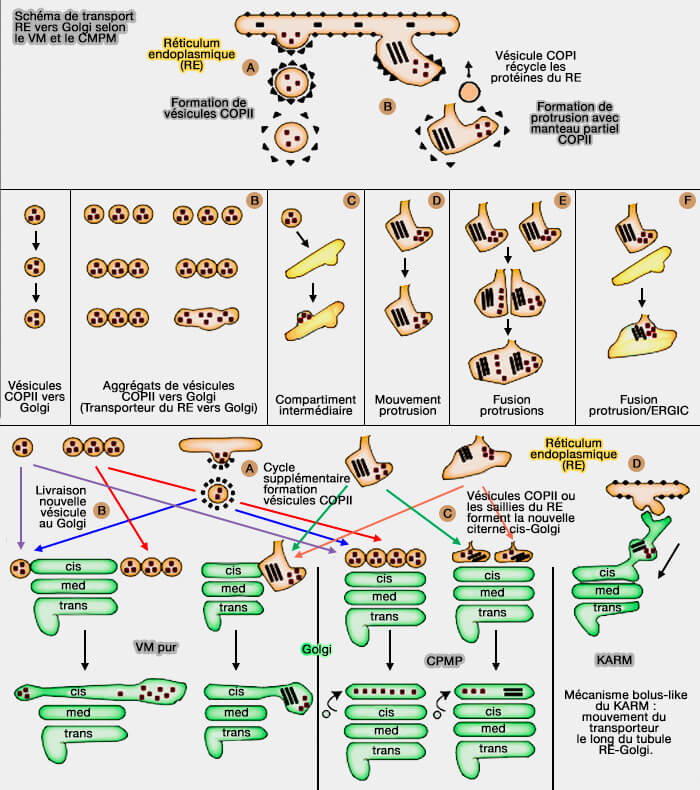
Modèle vésiculaire (VM)
Le modèle vésiculaire présume que des vésicules assurent le transport.
1. Des vésicules recouvertes d'un manteau de COPII bourgeonnent du réticulum endoplasmique (RE) et forment un compartiment intermédiaire avec l'appareil de Golgi appelé ERGIC (Endoplasmic-Reticulum–Golgi Intermediate Compartment, appelé aussi VTC).
2. Les protéines de l'ERGIC se déplacent vers le cis-Golgi par la formation de vésicules COPII qui bourgeonnent entre chaque compartiment, i.e. ERGIC, cis-Golgi, Golgi médian, trans-Golgi (TGN), cytosol, membrane plasmique.
- Les saccules (ou citernes) dans ce modèle sont stationnaires et les protéines golgiennes (GGE) restent en place.
- Les vésicules sécrétoires transportent les protéines du RE.
- Le transport vésiculaire est antérograde, RE vers cis-Golgi.
3. Toutefois, certaines observations contredisent le VM, comme celles sur les mégacargos ( VM de l'EGT et VM de l'IGT).
VM de l'EGT et VM de l'IGT).
(Figure : vetopsy.fr d'après Mironov et Beznoussenko)
Modèle classique de maturation-progression des citernes (CMPM)
1. Le modèle classique de maturation-progression des citernes (CMPM) suppose que :
- les transporteurs immatures du réticulum endoplasmique (RE) vers l'appareil de Golgi sont formés par la protrusion du RE,
- les citernes du Golgi mûrissent et se déplacent du cis-Golgi au trans-Golgi (TGN), puis vers la membrane plasmique où elles délivrent les protéines.
2. Toutefois, certaines observations contredisent le CMPM ( CMPM de l'EGT et CMPM de l'IGT).
- l'augmentation des concentrations (augmentation de la densité numérique) de tout cargo, et notamment des mégacargos,
- la vitesse de livraison du cargo du côté cis au côté trans du Golgi devrait être égale.,
- l'épuisement de quelques-unes des protéines résidentes des vésicules COPI.
Modèle de diffusion (DM)
1. Le modèle de diffusion (DM) implique, comme son nom l'indique, une diffusion le long de liaisons constantes entre le réticulum endoplasmique (RE) et l'appareil de Golgi.
- Si les citernes de Golgi sont bien connectées entre elles, aucune preuve n'a pu montrer une telle diffusion entre le RE et le Golgi.
- Ce modèle coexisterait avec le modèle classique de maturation-progression des citernes (CMPM) au niveau du Golgi pour les mégacargos comme le protocollagène.
2. Toutefois, certaines observations contredisent le DM ( DM de l'EGT et DM de l'IGT) :
- la concentration des cargos diffusibles,
- l'importance des protéines SNARE à chaque étape du transport, sans justification car il n'y a ni fusion ni fission,
- la rareté des connections,
- la présence d'empilements de saccules du Golgi sans connexions pendant le transport (Secretory traffic triggers the formation of tubular continuities across Golgi sub-compartments 2004),
- les concentrations croissantes des cargos, y compris les mégacargos pendant l'EGT et l'IGT.
Modèle KARM (Kiss-And-Run Model)
1. Pour résoudre les contradictions des modèles précédents, le modèle KARM (Kiss-And-Run Model) est un modèle qui est utilisé dans un bon nombre de processus comme (The Kiss-and-Run Model of Intra-Golgi Transport 2012) :
- le comportement des vésicules synaptiques (Synaptic vesicles: is kissing a matter of competence? 2001 et Synaptic Vesicle Exocytosis 2011),
- la libération des hydrolases des lysosomes vers les endo-lysosomes, les phagosomes, les autolysosomes ou les mélanosomes.
Le kiss-and-run est étudié dans un chapitre spécial.
(Figure : vetopsy.fr d'après Mironov et Beznoussenko)
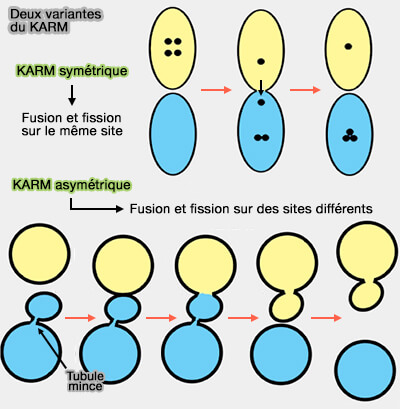
2. Le principe de base de KARM est que la fusion précède la fission, et même que cette fusion entraîne la fission (The golgi apparatus and centriole 2019).
a. La fusion/fission peut se produire sur un même site, i.e. variante symétrique du KARM.
Dans le cadre du KARM symétrique, le mécanisme de concentration implique des tubules étroits et une asymétrie de la composition ionique.
- Toutefois, des connections relativement épaisses peuvent induire une augmentation des concentrations de cargos dans l'un des deux compartiments.
- Si la délivrance des protons est asymétrique, la diffusion des agrégats de protéines vers l'arrière, i.e. transport rétrograde, est plus lente que dans le sens antérograde.
- La fusion et la fission doivent avoir des mécanismes moléculaires pour leur réalisation.
b. La fusion/fission peut se produire sur des sites différents, i.e. variante asymétrique du KARM, appelée aussi modèle de maturation des transporteurs (CMM ou carrier-maturation model).
Dans le cadre du KARM asymétrique :
- la fusion se situerait entre les bords des compartiments proximaux et distaux,
- la fission se situerait quelque part dans le compartiment proximal où des rangées de pores ou de tubules minces devraient être localisées,
- les protéines SNARE devraient être concentrées sur les domaines de fret.
3. Toutefois, des observations peuvent contredire le KARM ( KARM de l'EGT et KARM de l'IGT).
- a. Les cargos membranaires et les mégacargos doivent être organisés en domaines. Les grands domaines de fret sont plus efficaces pour le transport que les vésicules.
- b. Les protéines SNARE devraient être concentrées sur les domaines de fret.
- c. Entre le domaine cargo et le compartiment proximal, il doit y avoir un ou des tubules à membrane mince, qui relient le domaine cargo aux domaines du compartiment proprement dit.
- d. Les pores doivent être consommés pendant le transport.
- e. Une ligne de régression exponentielle négative pour le processus de vidange du Golgi en soi devrait exister dans toutes les conditions.
- f. Pour le transport EGT et post-Golgi, le mécanisme de type bolus doit être optimal.
- g. Tous les compartiments doivent toujours être connectés, ce qui est contredit par l'existence de nombreux SNARE.

À l'heure actuelle, il existe plus de 20 variantes des 4 modèles principaux et les différents modèles pourraient ne pas être incompatibles (Models of Intracellular Transport: Pros and Cons 2019).
Retour au système endomembranaire
Biologie cellulaire et moléculaireConstituants de la celluleTransport membranaireTransports sans mouvements membranairesTransports passifsTransports actifsTransports avec mouvements membranaires : trafic vésiculaireFusion/fission membranaireEndocytoseVoie sécrétoireCanaux ioniquesTransporteursUniportsPompesCo-transporteursMoteurs moléculairesVoies de signalisation