Comme le modèle proposé est basé principalement sur des " instantanés " du domaine moteur dans les différents stades nucléotidiques en l'absence de microtubules, la séquence temporelle des changements conformationnels dans le grand domaine ATPase reste discutée.
2. On spécule sur la manière dont les événements dans le domaine ATPase (hydrolyse de l'ATP, libération de Pi de AAA1 et powerstroke) sont couplés à la liaison MTBD/microtubule, événements essentiels par le passage d'un stade de faible affinité à un stade de haute affinité des MTBD pour les microtubules.
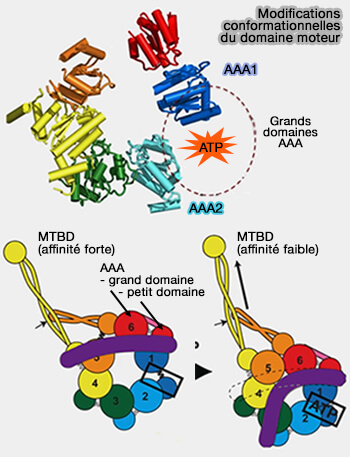
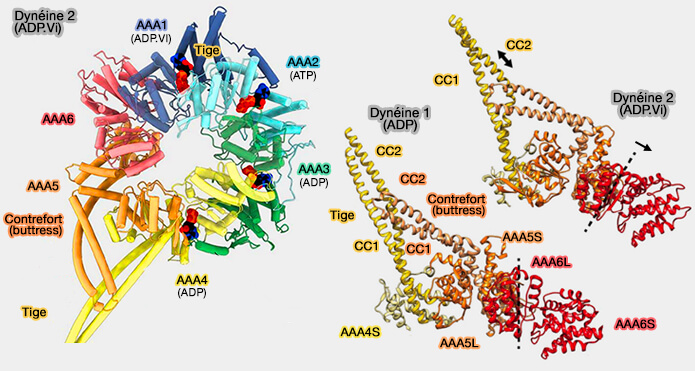
Domaine moteur, tige et contrefort de la dynéine
(Figure : vetopsy.fr d'après Roberts et Rao et coll)
a. Le contrefort s'éloigne de la tige, ce qui provoque un changement d'affinité des MTBD pour les microtubules.
Changements de registre des MTBD
(Figure : vetopsy.fr d'après Nishida et coll)
Ce changement est obtenue par des modifications dans le mode d'association du domaine coiled-coil (superhélice) appelé " registre " (registry), constitué de CC1 et CC2 lié au MTBD.
Le glissement de CC1 par rapport à CC2 d'un tour d'hélice α entraîne un changement de l'affinité pour les microtubules.
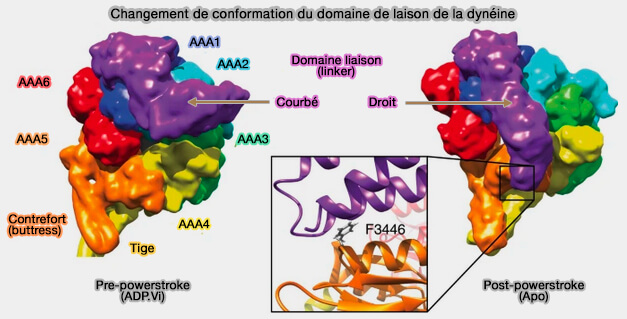
Dans le stade ADP-vanadate, correspondant au stade ATP, l'extrémité N-terminale du domaine de liaison plié est positionnée en AAA2/3.
b. Le domaine de liaison passe de la conformation droite à courbée, i.e. l'extrémité N-terminale du domaine de liaison plié est positionnée en AAA2/3. (recovery stroke ou course de récupération ou pré-powerstroke).
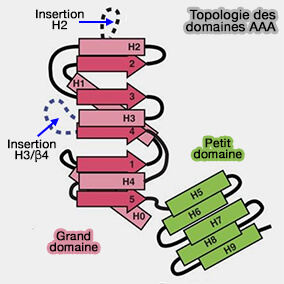
Certains éléments de la structure secondaire des domaines AAA1-5 , i.e. appelés " insertions ", insert en anglais, extensions α-hélicoïdales ou épingle à cheveux β, situées sur la face du domaine de liaison de l'anneau AAA+, pourraient être responsables de son remodelage.
L'insertion H2, épingle à cheveux β qui émerge de l'hélice H2 du domaine AAA, pointe vers le centre de l'anneau AAA+ pour interagir avec le substrat dans certaines protéines AAA+.
L'insertion pré-sensorielle I (PS-I) est une insertion entre l'hélice 3 et le brin 4 du domaine AAA. Il semblerait que l'insertion PS-I du domaine AAA4 provoquerait un conflit stérique avec l'extrémité N-terminale du linker lors de la fermeture de l'anneau AAA+. Induit par la présence d'ATP, elle déclencherait la flexion du domaine de liaison (How AAA+ ring opening and closing coordinates microtubule binding and linker movement 2015).
Topologie des AAA
(Figure : vetopsy.fr)
Les insertions PS-I et H2 du domaine AAA2 ont également été associées au remodelage du linker. Elles sembleraient interagir avec la région du point pivot dans le domaine de laison pour participer au remodelage de ce dernier (Crystal Structure of the Dynein Motor Domain 2011).
c. Le domaine moteur recherche un nouveau site de liaison sur le microtubule, via un mode d'interaction de faible affinité.
Le mouvement brownien, i.e. collision entre les molécules d'eau et la dynéine, qui provoque le tremblement des molécules de dynéine peut faciliter cette recherche
2. Comment expliquer ce phénomène ?
a. La liaison de l'ATP en AA1 modifie la rotation entre les grands et les petits domaines d'une sous-unité ( structure des AAA+ ATPases)
Or, ce site de liaison de l'ATP, la poche nucléotidique, se retrouve entre le petit domaine d'une unité et le grand domaine de l'unité suivante, i.e. la liaison du nucléotide referme l'espace entre AA1 et AA2, ce qui permet :
Quand l'ATP est fixé sur AA1 et sur AA3, le domaine de liaison ne se courbe pas, i.e. les changements conformationnels de la tige ne se produisent pas.
Les stades de haute et faible affinité aux microtubules sont régulés par l'introduction de ponts disulfure dans la tige.
Conformation des MTBD
Le MTBD de la dynéine peut prendre trois conformations majeures :
le registre +β,
le registre semi-α,
le registre α.
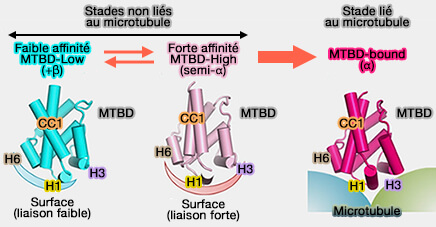
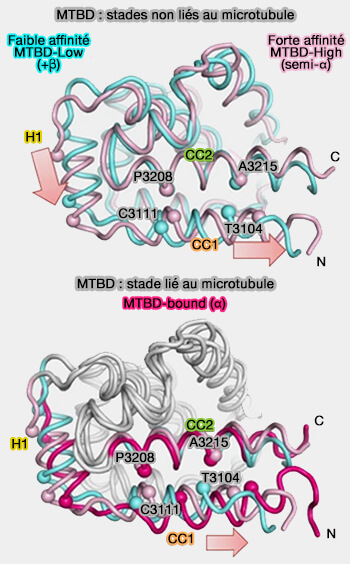
États majeurs des MTBD
(Figure : vetopsy.fr d'après Nishida et coll)
1. Dans les stades non liés au microtubule, le MTBD est en équilibre conformationnel entre MTBD-Low (registre +β) et MTBD-High (registre semi-α).
Dans le stade de faible affinité du MTBD (MTB-Low), MTBD adopte un registre +β, i.e. les régions H1 et H6 formant une surface de liaison faible pour les microtubules.
Dans le stade de forte affinité du MTBD (MTB-High), MTBD adopte un registre semi-α avec glissement de CC1 d'un demi-tour de l'hélice α, les régions H1, H3 et H6 formant une surface à forte liaison.
2. Dans le stade lié au microtubule, le MTBD subit un autre changement de conformation pour adopter le registre α en glissant d'un tour d'une hélice α au total.
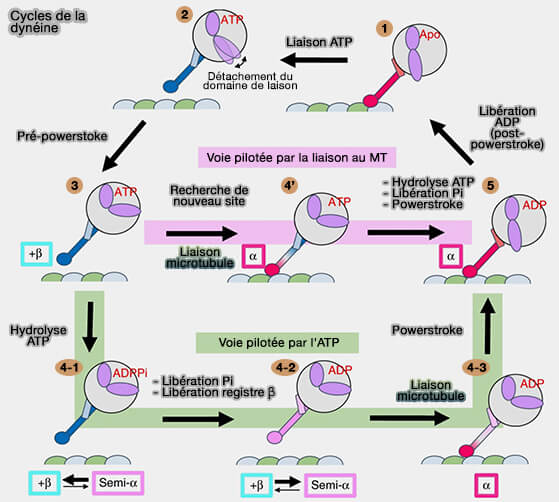
Cycles de la dynéine
Voie pilotée par l'ATP
1. Dans la voie pilotée par l'ATPase (vert), l'hydrolyse de l'ATP (3 à 4-1) et la libération ultérieure de Pi (4-1 à 4-2) déplacent l'équilibre du MTBD vers le registre semi-α (4-2) pour augmenter l'affinité de liaison à la MT.
2. Après la liaison au microtubule, le registre α stabilisé transmet les changements conformationnels au domaine ATPase, facilitant ainsi le powerstroke du domaine de liaison (4-3 à 5).
Cycles de la dynéine en fonction des MTBD
(Figure : vetopsy.fr d'après Nishida et coll)
Voie pilotée par par la liaison au microtubule
1. Dans la voie induite par la liaison au microtubule (rose), la dynéine liée à l'ATP recherche un nouveau site de liaison dans l'état de faible affinité (3).
2. Lors de la liaison de la MT (4') , la stabilisation du registre α transmet les changements conformationnels au domaine ATPase, ce qui facilite l'hydrolyse de l'ATP, la libération de Pi et le powerstroke en même temps (5).
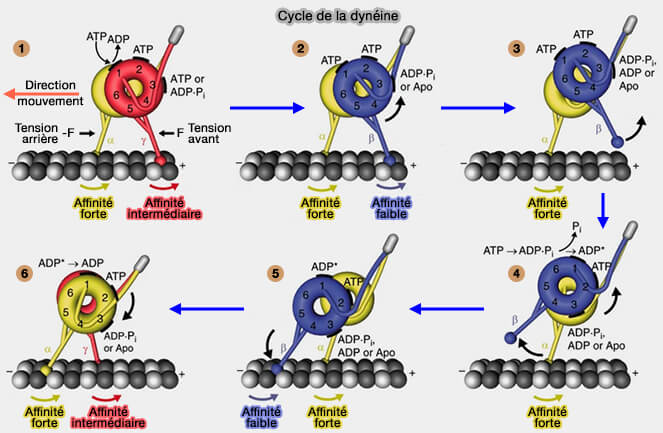
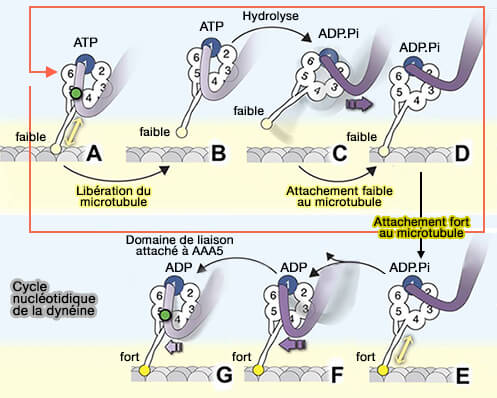
1. Après la libération de l'ADP par AAA1, on assiste à la liaison de l'ATP sur AA2 et AA3.
2. La liaison de l'ATP sur AA1 provoque :
la libération de AAA5 du domaine de liaison,
la transition du registre γ des hélices de tige avec une faible affinité MT intermédiaire vers le registre β faiblement lié à la MT.
Cycle de la dynéine
(Figure : vetopsy.fr d'après Rao et coll)
3. Le détachement de la tête arrière se produit lorsque AAA3 est dans l'état de transition ADP∙Pi ou lié à l'ADP.
4. L'ATP de AA1 est à hydrolysé et on assiste à la libération à la libération de Pi et le " réarmement " du domaine de liaison (priming stroke ou coup d'amorçage) qui déplace la tête détachée vers un nouveau site du microtubule tandis que la tête attachée au microtubule supporte la charge.
5. On observe une nouvelle liaison au microtubule dans le registre β faiblement lié au microtubule et la transition de l'état " ADP* à haute énergie " à l'état " ADP à basse énergie ".
6. Cette transition génère un retour du domaine de liaison (powerstroke), entraînant son amarrage du domaine de liaison à AAA5 et la transition vers le registre α fort de liaison MT des hélices de tige.
Une condition préalable à l'état de liaison forte est que AAA3 ne soit pas lié à l'ADP, ce qui suggère que AAA3 est toujours dans l'état ADP∙Pi ou sans nucléotide.




 structure des AAA+ ATPases)
structure des AAA+ ATPases)