L'ATP synthase ou F1FO-ATPase est composée, outre la région catalytique F1, par un rotor formé par la région FO qui correspond à une pompe à proton (H+).
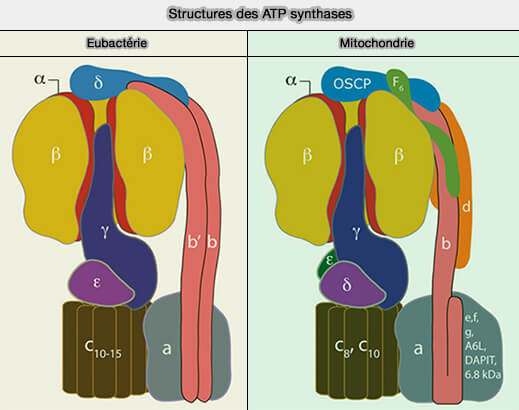
Structures des ATP synthases
(Figure : vetopsy.fr d'après Walker)
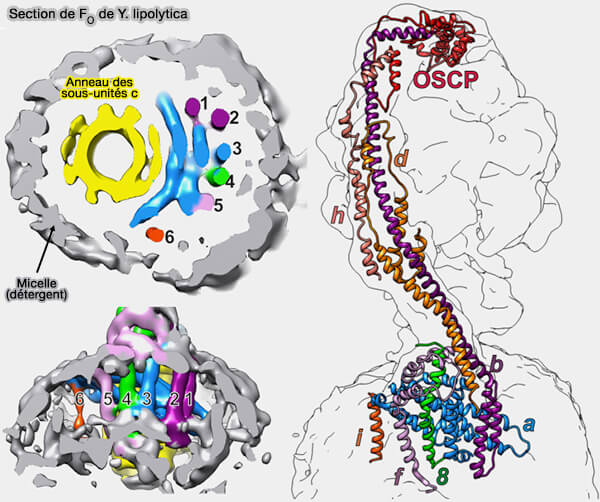
Structure générale de la région FO de l'ATP synthase
La région FO, O pour oligomycine et non pas le chiffre 0, désignée ainsi car cet antibiotique cible cette fraction, est une protéine insoluble qui possède plusieurs sous-unités différentes :
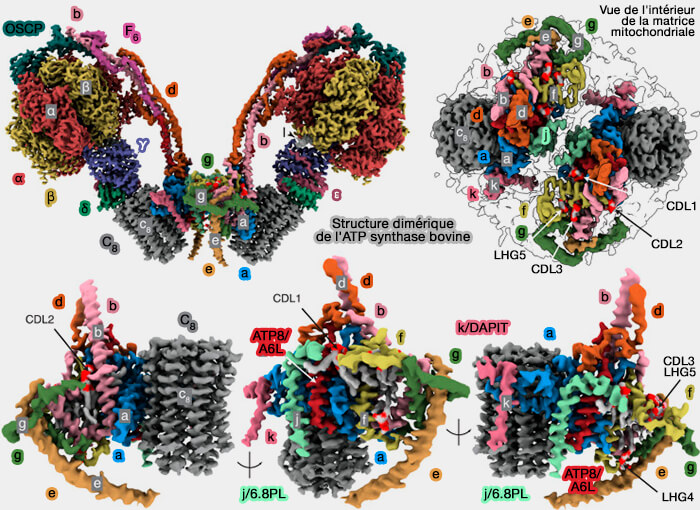
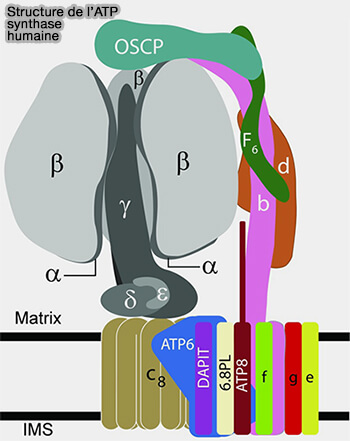
Structure de l'ATP synthase humaine
(Figure : vetopsy.fr d'après He et coll)
c, a et b chez les bactéries,
c, a/ATP6, ATP8/A6L, e, f, g, k/DAPIT (diabetes-associated protein in insulin-sensitive tissue) et j/6.8PL (6.8-kDa proteolipid) chez les eucaryotes.
Remarque : les sous-unités F6, b et d mitochondriales, i.e. b et b' des bactéries et des chloroplastes, peuvent être considérées comme des sous-unités de FO.
FO est mobile dans le plan de la membrane, et peut tourner autour d’un axe constitué par certaines des sous-unités de F1, d’où sa qualification de rotor/essieu.
2. Par opposition à la région F1 qui est le stator, FO est qualifiée de rotor.
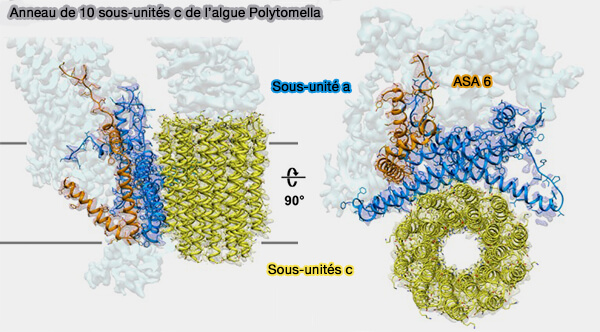
Anneau de sous-unités c
Structure d'ensemble
1. Les sous-unités c, en nombre variable selon les espèces, forme un anneau qui passe par des changements conformationnels lorsqu'elles sont protonées et déprotonées ( mécanisme de FO).
Ce nombre varie : 8 dans les mitochondries bovines, 10 dans les mitochondries de levure, 14 dans les chloroplastes et de 10 à 15 chez les bactéries.
On pense que le nombre de c est déterminé par le rapport ion/ATP, qui se situe entre 2,67 pour 8 c et 5,00 pour 15 c.
Les organismes, comme les mammifères, qui possèdent une force motrice des protons (PMF ou Δp) à haute vitesse constante, n'ont besoin que d'un nombre faible de c, alors que ceux exposés à des facteurs de traitement variables comme les chloroplastes et/ou à des PMF à faible taux comme chez les bactéries ont besoin d'un nombre plus élevé de c.
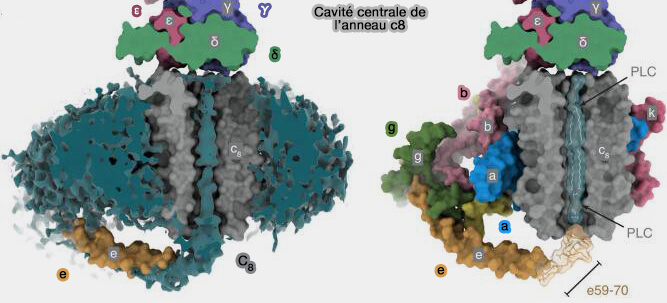
La moitié inférieure a été attribuée à un autre lipide mobile et à la région C-terminale de la sous-unité e, qui s'étend vers la cavité centrale dans l'espace intermembranaire et pourrait simplement pénétrer dans la cavité chez les bovins, mais pas, semble-t-il, chez la levure (Atomic model for the dimeric FO region of mitochondrial ATP synthase 2019).
Cavité centrale de l'anneau c8 de l'ATP synthase bovine
(Figure : vetopsy.fr d'après Spikes et coll)
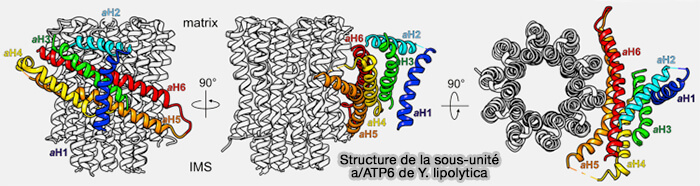
aH3-aH6 forme un faisceau hélicoïdal, i.e. aH5 contiendrait Arg182, qui interagit avec le glutamate de l’anneau c protonatable.
La sous-unité a est essentielle au mécanisme de translocation des protons par ces deux demi-canaux ( translocation des protons).
Structure de la sous-unité a/ATP6 de Y. lipolytica
(Figure : vetopsy.fr d'après Hahn et coll)
2. La sous-unité b a deux α-hélices transmembranaires N-terminales interagissant directement ou indirectement avec les sous-unités e, f, g, k/DAPIT et j/6,8PL, qui n’ont aucun rôle connu dans la synthèse ou l’hydrolyse de l’ATP.
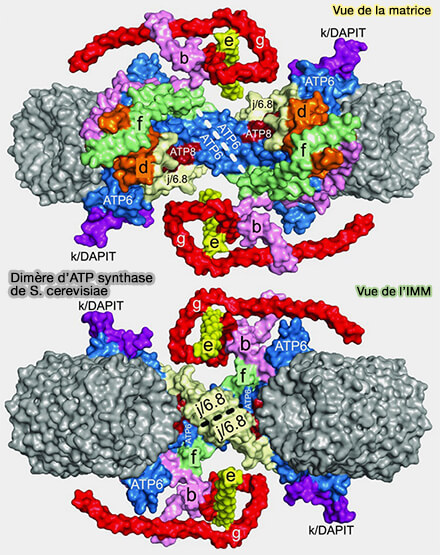
Dimère d’ATP synthase de S. cerevisiae
(Figure : vetopsy.fr d'après He et coll)
Le domaine membranaire de la sous-unité b est associé aux sous-unités a/ATP6 et ATP8/A6L codées par les mitochondries.
La région N-terminale d’ATP8 a une seule hélice α transmembranaire, et sa région C-terminale s’étend dans la tige périphérique.
a. L'ATP8 et la sous-unité b maintiennent l’ATP6 en contact avec l’anneau c.
b. La sous-unité b relie FO au sommet de l'hexagone (αβ)3 de F1.
3. D'autres sous-unités sont étudiées dans la dimérisation.
une hélice transmembranaire N-terminale avec un motif GxxxG essentiel conservé, une signature d’interaction hélice-hélice
une extrémité C-terminale hydrophile qui expliquerait son attache à l’IMS.
b. La sous-unité g comprendrait :
un domaine matriciel N-terminal,
une hélice transmembranaire C-terminale qui contient également un motif GxxxG conservé.
La sous-unité g pourrait être liée à l’extrémité N-terminale de b dans la matrice car la suppression de la première hélice transmembranaire de b entraîne la perte de g et la dissociation du dimère.
c. Les sous-unités e et g peuvent ainsi former un hétérodimère serré dans la membrane via leurs motifs GxxxG.
Cavité centrale de l'anneau c8 de l'ATP synthase bovine
(Figure : vetopsy.fr d'après Spikes et coll)
Chez les mammifères
La dimérisation dans de l'ATP synthase des mammifères est plus complexe.
La sous-unité f stabilise les dimères de l’ATP synthase, mais pas les monomères.
L’activité synthétique/hydrolytique de l’ATP synthase n’est pas altérée dans les KO de la sous-unité f.
La régulation négative de la sous-unité f humaine perturbe la morphologie normale des crêtes mitochondriales.
La sous-unité f module la taille et la sensibilité du pore de transition de perméabilité.
2. DAPIT (diabetes-associated protein in insulin-sensitive tissue) , appelée aussi k comme l'orthologue de la levure, pourrait être impliquée dans la formation de liens entre les dimères dans les rangées de dimères des crêtes.
3. La sous-unité e, mal connue, interviendrait aussi dans la dimérisation.





 mécanisme de FO).
mécanisme de FO).