Transport membranaire
Transports sans mouvements membranaires
Transporteurs actifs : co-transporteurs (transporteurs secondaires)
AE1 ou Band 3 (Bande 3)
Mécanisme alternatif et fonctions
AE1/Band 3, antiport (co-transporteur) hautement conservé, fait partie des échangeurs anioniques qui régulent l'échange Cl- /HCO3- (chlore/bicarbonate) à travers les membranes plasmiques (transport du bicarbonate par les SLC4).
Fonctionnement de AE1/Band 3
1. Le mouvement de bascule relatif du domaine central et du domaine de la porte dans ces co-transporteurs modifie l'accessibilité du seul site de liaison au substrat d'un côté de la membrane à l'autre pour accomplir la translocation anionique (Solute carriers keep on rockin' 2015).
La plupart des co-transporteurs peuvent être décrits comme fonctionnant sous :
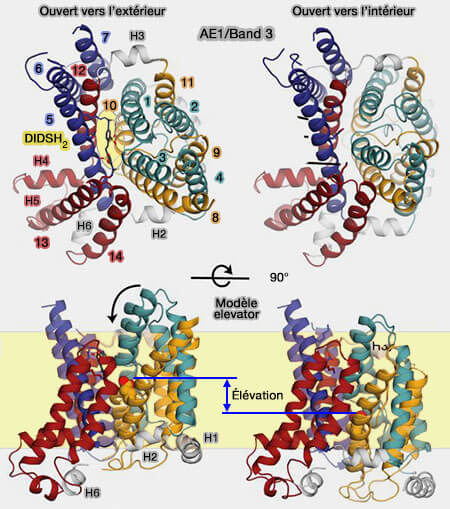
3. La protéine oscille entre deux états conformationnels majeurs qui exposent le site de liaison anionique :
à l'espace extracellulaire (OF, outward-facing) comme dans le cas de la liaison avec DIDSH2,
à l'espace intracellulaire (IF, inward-facing), en l'absence d'anion, ce qui est ici son étape limitante, mais pas aux deux simultanément.
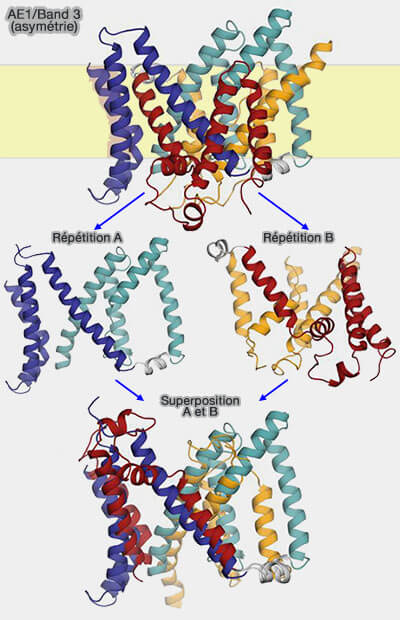
a. Band 3 est formée de deux répétitions inversées (structure).
Toutefois, l'orientation de TM1-TM4 par rapport à TM5-TM7 diffère de celle de TM8-TM11 par rapport à TM12-TM14, i.e. les répétitions A et B n'ont pas la même structure.
Cette différence structurelle est en effet cruciale, car c'est la raison pour laquelle le site supposé de liaison aux anions est exposé à l'espace extracellulaire.
Dans la configuration OF (liée au DIDSH2), le site présumé de liaison au substrat est situé entre les extrémités des parties hélicoïdales de TM3 et TM10 dans le domaine de transport et est accessible du côté extracellulaire de la membrane.
Dans la configuration IF (modèle à répétition permutée), le site supposé de liaison aux anions est exposé au cytoplasme et non au côté extracellulaire.
Modèle elevator de Band 3
(Figure : vetopsy.fr d'après Ficici et coll)
la préservation de la structure secondaire de la protéine,
la préservation de la structure tertiaire individuelle des domaines centraux et des portes.
4. Cette transition nécessiterait :
aucun mouvement à l'interface de dimérisation, i.e. elle est stationnaire ;
une translation du site de liaison des anions (TM1/TM8 et TM3/TM10) d'environ 11 Å (deux tours d'hélice) et une rotation d'environ 17 ° autour d'un axe perpendiculaire à la membrane et centré sur l'interface du dimère.
a. La translation résultante serait de 8 Å, suggérant que AE1 utilise un mécanisme elevator pour exposer alternativement le site de liaison d'anion de chaque côté de la membrane.
b. Ce changement est très rapide : AE1 est l'échangeur le plus rapide connu.
Ce mécanisme nécessite une faible affinité pour les anions qui doivent se délier très vite.
En effet, l'anion est lié, non par de multiples chaînes latérales basiques, sauf peut-être Arg730, mais plutôt faiblement entre deux dipôles hélicaux (TM3 et TM10), qui plus est avec une charge négative voisine fournie par Glu681, agissant comme une porte pour déplacer les anions liés.
Par contre, l'uracile liée dans UraA est maintenue en place par de multiples liaisons qui doivent être rompues et recrées à chaque cycle, d'où un transport beaucoup plus lent.
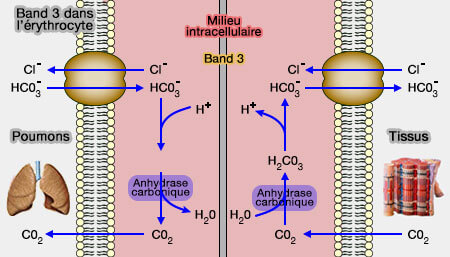
AE1 transporte du bicarbonate (HCO3-), formé à partir de dioxyde de carbone, en échange de chlorure (Cl-).
Band 3 dans l'érythrocyte
(Figure : vetopsy.fr)
Ce mécanisme diminue le pH à l'intérieur des cellules sanguines, de sorte que l'oxygène est libéré de l'hémoglobine et peut diffuser dans les tissus.
1. Le dioxyde de carbone (CO2), généré par les processus métaboliques tissulaires, diffuse dans les globules rouges.
3. L'augmentation de la concentration de bicarbonate dans l'érythrocyte provoque leur transport par AE1 dans le plasma sanguin par un échange électroniquement neutre avec les ions chlorure (Cl-).
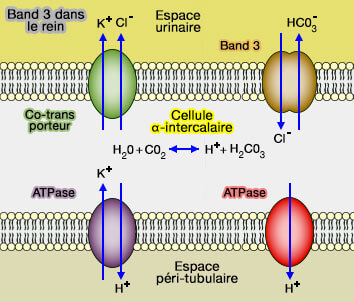
Band 3 dans le rein
(Figure : vetopsy.fr)
Le transport est extrêmement rapide, le plus rapide connu, car le nombre de transporteurs est élevé (106 AE1/cellule), chacun échangeant 4 à 5 x 104 ions/seconde : 90 % du CO2 des tissus est envoyé vers les poumons sous cette forme soluble.
4. Dans le globule rouge, cette réaction provoque l'accumulation de protons, donc une diminution du pH intra-érythrocytaire.
5. Par ce faible pH, l'hémoglobine (oxyhémoglobine) libère son l'oxygène qui diffuse dans les tissus.
6. Lorsque le sang atteint les poumons, le processus est inversé et le CO2 est exhalé.
Dans le rein, Band 3 est localisée sur la surface basolatérale des cellules intercalaires A du canal collecteur du rein, cellules qui acidifient l'urine en sécrétant des protons (H+) par des ATPases.




 transport du bicarbonate par les SLC4).
transport du bicarbonate par les SLC4).