Transport membranaire
Transports sans mouvements membranaires
Transporteurs actifs : co-transporteurs (transporteurs secondaires)
Mécanismes de transport : vue d'ensemble
1. Les pompes (ou transporteurs primaires)utilisent l'énergie de l'hydrolyse de l'ATP (adénosine triphosphate) pour faire traverser la membrane plasmique à la molécule.
Quand on utilise le terme de transporteurs dans les publications, on parle le plus souvent des co-transporteurs ou transporteurs secondaires.
Vue d'ensemble du mécanisme
Un mécanisme simple pour faire passer la membrane à des ions et/ou à des molécules est d'exposer alternativement un site de liaison au substrat de chaque côté de la membrane.
Son modèle d'accès alternatif a été depuis largement affiné par les avancées sur les structures des transporteurs et on reconnaît, à l'heure actuelle, trois modèles de transport.
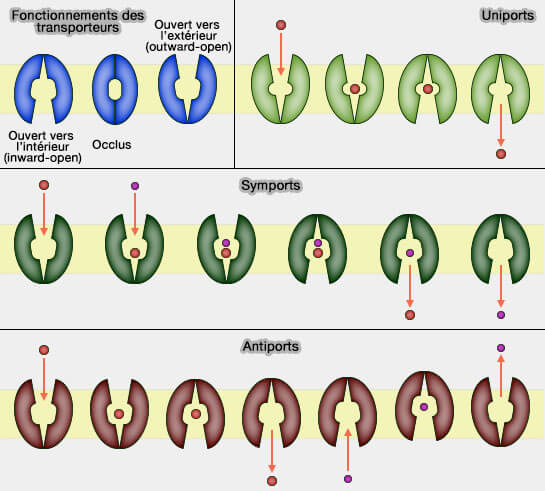
Types des transporteurs
(Figure : vetopsy.fr)
Ce processus implique :
1. deux portes, une extramembranaire et l'autre intramembranaire, qui ne peuvent pas être ouvertes en même temps ;
2. une conformation ouverte vers l'extérieur (outward-facing ou outward-open) ;
3. une conformation ouverte vers l'intérieur (inward-facing ou inward-open) ;
4. un changement conformationnel du transporteur qui passe par une étape où les deux portes sont fermées (conformation occluse), excepté peut-être pour le mécanisme le plus simple des rocker-switch.
de deux solutés ou plus dans la même direction (symport) : le changement conformationnel vers l'extérieur et vers l'intérieur ne peut pas se produire lorsqu'un seul substrat est lié,
deux solutés dans des directions opposées (antiport ou échangeur) : le changement conformationnel vers l'extérieur et vers l'intérieur ne peut se produire que lorsque l'un des substrats est lié.
Déductions
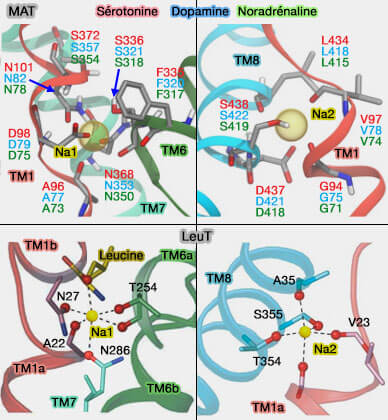
Couplage du substrat et des ions
L'aspect énergétique clé du mécanisme est que le substrat et les ions conducteurs sont transportés ensemble, jamais seuls.
1. Dans les symports, le couplage peut être réalisé pendant la liaison, pendant la translocation de substrat ou pendant les deux.
La liaison du substrat et des ions doit être coopérative, i.e. aucun des deux ne se lie efficacement seul.
Même si cette liaison se produit indépendamment, la liaison des deux est nécessaire pour le changement conformationnel du co-transporteur.
2. Dans les antiports, le couplage des ions est nécessaire pour la libération du substrat, pour le retour du transporteur à sa conformation de départ, ou pour les deux.
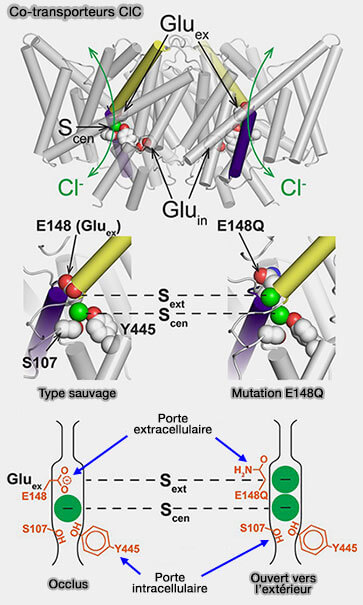
Accessibilité alternative
aux sites de liaison
Le site du substrat doit être accessible de manière alternative, et ce d'un seul côté de la membrane :
soit dans l'espace extramembranaire (OF, outward-facing ou outward-open),
soit dans l'espace intramembranaire (IF, inward-facing ou inward-open).
Cette accessibilité alternée ne peut être obtenue que par le couplage allostérique des portes extracellulaire et intracellulaire du transporteur, i.e. les transitions structurelles globales sont nécessaires à la translocation du substrat et des ions.
Plusieurs mécanismes de transport alternatifs ont été décrits et la plupart des transporteurs peuvent être décrits comme fonctionnant selon 3 modèles.
1. Dans deux modèles,
le site de liaison au substrat est situé à l'interface de deux domaines, approximativement à mi-chemin de la membrane ;
la barrière entre les deux domaines s'écarte du côté intracytoplasmique pour se reformer du côté extracellulaire, i.e. la barrière est mobile.
Le substrat, qui se lie entre les deux domaines, induit un changement conformationnel du transporteur autour de ce site de liaison.
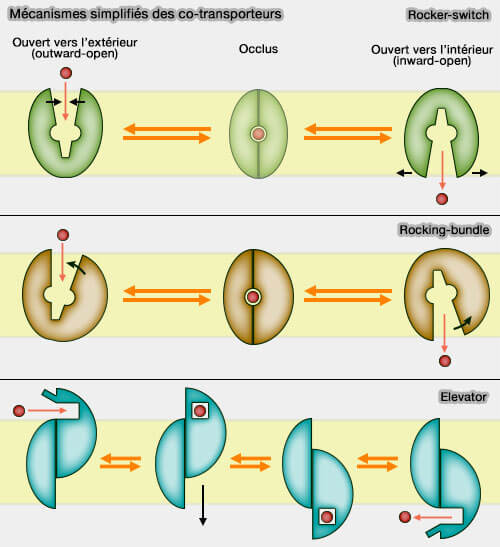
Mécanismes simplifiés des co-transporteurs
(Figure : vetopsy.fr) Dans le modèle rocker-switch (" commutateur ou interrupteur à bascule "), le plus simple, les deux domaines sont similaires structurellement et basculent comme un interrupteur : c'est le cas des SemiSWEET, des EmrE et des PnucC, mais surtout de la plus grande superfamille des transporteurs secondaires, les transporteurs MFS (Major Facilitator Superfamily).
2. Le modèle elevator (" ascenseur ou élévateur" ) a été décrit plus récemment.
Le site de liaison au substrat se trouve en grande partie ou en totalité dans un seul domaine relativement rigide.
Ce domaine de transport se déplace à la verticale (mais aussi, avec une légère rotation) contre le domaine d'échafaudage qui reste ancré, presque immobile, dans la membrane.
La barrière reste ainsi dans une position fixe, contrairement aux autres modèles.






 autres transporteurs).
autres transporteurs).