Transport membranaire
Transports sans mouvements membranaires
Transporteurs actifs : co-transporteurs (transporteurs secondaires)
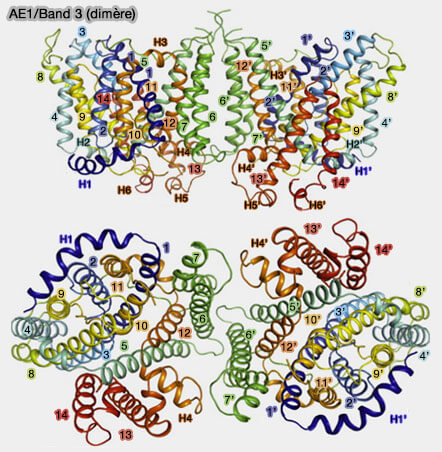
AE1 ou Band 3 (bande 3)
Vue d'ensemble et structure
Band 3 (bande 3 ou AE1) fait partie des échangeurs anioniques (antiports faisant partie des co-transporteurs ou transporteur secondaires) qui régulent l'échange Cl-/HCO3- (chlore/bicarbonate) à travers les membranes plasmiques.
Band 3 (Band 3 anion transport protein ou protéine érythrocytaire-1 échangeuse d'anions en français) est aussi appelé AE1 (anion exchanger 1) ou SLC4A1 (solute carrier family 4 member).
1. AE1/Band 3 (omim.org), omniprésente chez les vertébrés, est exprimée chez les mammifères dans :
la membrane cellulaire des érythrocytes (globules rouges) par son isoforme eAE1, dans laquelle elle représente la principale glycoprotéine (25 à 30 %),
la surface basolatérale des cellules intercalaires A du canal collecteur du rein, cellules qui acidifient l'urine en sécrétant des protons (H+) par des ATPases, par son isoforme kAE1 (k pour kidney),
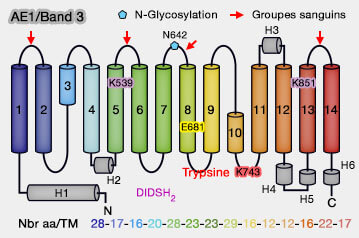
Le domaine membranaire (361-911), dont l'extrémité intracytoplasmique est désordonnée, comprend 14 domaines transmembranaires (TM1-TM14).
Ce domaine, impliqué dans l'échange d'ions, est fonctionnel en l'absence du domaine N-terminal intracytoplasmique. La partie hautement conservée commence à Phe379.
1. Les domaines transmembranaires ont une conformation hélicoïdale classique.
Certains n'atteignent pas les 20 résidus, correspondant à l'épaisseur d'environ 30 Å de la phase hydrophobe d'une bicouche lipidique.
D'autres sont classées en boucles réentrantes (ou demi-hélices) comme TM3 ou TM10.
2. Les antigènes des groupes sanguins sont situés sur les boucles extracellulaires reliant les segments TM.
3. Le site unique de N-glycosylation sur Asn642 se trouve entre TM7 et TM8.
Organisation des domaines
transmembranaires
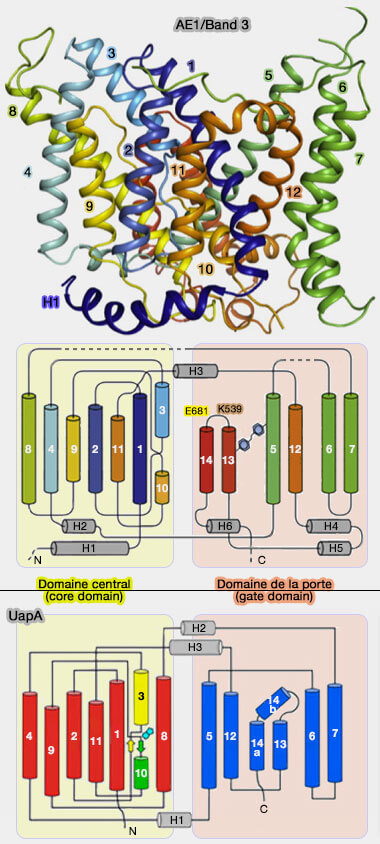
Les 14 segments TM de la bande 3 sont constitués de deux régions répétées (A et B) de 7 TM inversées (TM1-7 et TM8-14) à repliement similaire :
le domaine central (core domain ou domaine de transport), formé par TM1-4 et de sa répétition inversée TM 8-11 ;
le domaine de la porte (gate domain), constitué de TM5-7 étroitement associé à la répétition inversée TM12-14.
1. Par contre, l'orientation de TM1-TM4 par rapport à TM5-TM7 diffère de celle de TM8-TM11 par rapport à TM12-TM14, i.e. les répétitions A et B n'ont pas la même structure même si elles ont des topologies identiques (mais inversées).
Cette différence structurelle est en effet cruciale, car c'est la raison pour laquelle le site supposé de liaison aux anions est exposé à l'espace extracellulaire.
2. Le mouvement de bascule relatif des domaines de la porte et du noyau dans ces protéines modifierait l'accessibilité du seul site de liaison au substrat d'un côté de la membrane à l'autre pour accomplir la translocation anionique (mécanisme alternatif).
Site de liaison des anions
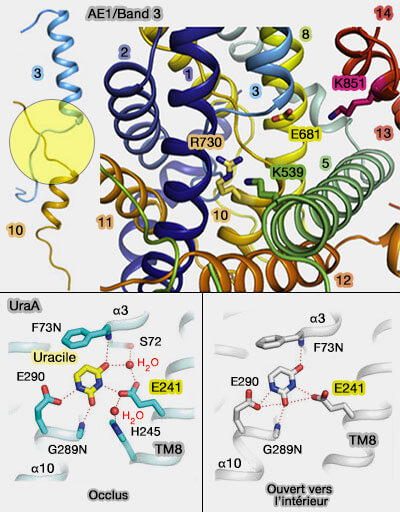
1. Le site de liaison aux anions est situé à l'extrémité N-terminale des hélices TM3 et TM10.
La charge négative de Glu681 pourrait empêcher la liaison des anions divalents comme le sulfate (SO4--) en raison de la répulsion ionique, tandis que l'élimination de la charge sur Glu681 permettrait la liaison au sulfate.
3. La chaîne latérale de Arg730 de TM10 fait face au site actif, situé à 7,5 Å de Glu681.
Arg730 fournit donc l'environnement chargé positivement pour accueillir les anions.
Arg730 peut interagir directement avec la charge négative sur le bicarbonate (et le chlorure), le groupe OH du bicarbonate formant une liaison hydrogène directement ou par l'intermédiaire de l'eau avec la chaîne latérale de Glu681.
Des résidus hydroxyles dans les régions étendues de TM3 (Ser465) et TM10 (Ser725, Thr727, Thr528) pourraient participer à la liaison des anions en remplaçant les molécules d'eau liées : les chlorures pourraient alors être déshydratés comme dans les canaux ClC (ClC chloride channels viewed through a transporter lens 2006).
Domaine de la porte (gate domain)
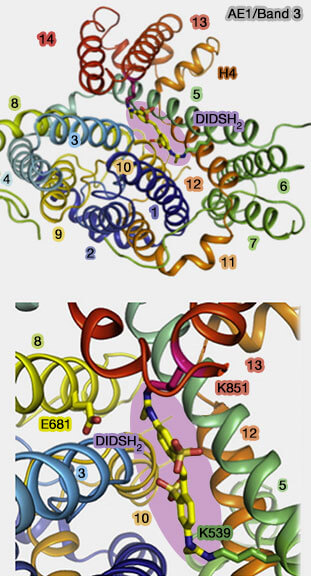
Le domaine de la porte contient des lysines dans TM5 (Lys539) et TM13 (Lys851).
1. Or, Le transport d'anions dans les globules rouges peut être inhibé par des anions organiques comme le 4,4'-diisothiocyano-2,2'-stilbène disulfonate (DIDS) ou son dérivé réduit (DIDSH2).
DIDSH2 se lie à ces lysines pour obstruer l'espace entre les lysines.
En outre, il interagit avec Arg730 qui fait face à l'espace entre les extrémités hélicoïdales de TM3 et TM10.
Il pourrait aussi entrer en interaction avec Glu681.
2. DIDSH2 occupe une position stratégique, entre le domaine central et les domaines de la porte, dans un passage qui mène au site de liaison des anions : il entraverait le mouvement relatif des deux domaines, bloquant la translocation en verrouillant Band 3 dans un état orienté vers l'extérieur (OF, outward-facing).
Domaine de dimérisation membranaire
Band 3 est présente sous forme dimérique et tétramérique dans les membranes.
a. L'interface dimérique (1092 Å2) est représentée par :
un faisceau à quatre hélices provenant de TM5 et TM6 dont les interactions prédominantes se produisent aux extrémités extracellulaires de ces deux hélices à Leu572 située à l'extrémité extracellulaire N-terminale de TM6 ;
une contribution de TM7 et du côté membranaire cytosolique de H4 et de la boucle le reliant à TM13.
b. Cette interface a une forme d'entonnoir, avec peu d'interactions entre les deux sous-unités sur le côté cytosolique du dimère.
L'interface dimère, constituée uniquement par le domaine de la porte, peut donc changer pendant le cycle de transport lorsque les sous-unités constituantes passent d'une conformation vers l'extérieur à une conformation tournée vers l'intérieur.
L'interface dimérique peut rester fixe et le domaine central peut se déplacer pendant le cycle de translocation.
Certains pensent que la dimérisation pourrait aussi avoir lieu par une interface entre les domaines centraux (Solute carriers keep on rockin' 2015).




 chapitre spécial), et plus particulièrement aux SLC4 (Solute Carrier Family SLC41, what do we really know about it? 2014).
chapitre spécial), et plus particulièrement aux SLC4 (Solute Carrier Family SLC41, what do we really know about it? 2014).