Transport membranaire
Transports sans mouvements membranaires
Transporteurs actifs : co-transporteurs (transporteurs secondaires)
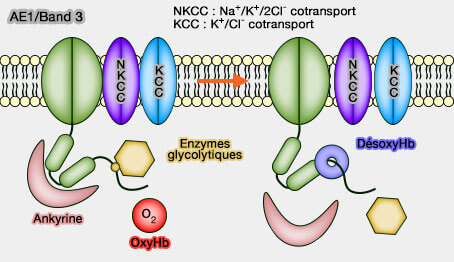
AE1 ou Band 3 (Bande 3)
Interactions : complexes membranaires et hémoglobine
Le domaine d'interaction N-terminal de Band 3 ou AE1, domaine de dimérisation (comme le domaine membranaire), comprend plusieurs sites de liaison qui permettent la formation de complexes fonctionnels essentiels à la cellule.
2. un domaine d'interaction, point d'ancrage pour de nombreuses protéines :
à l'ankyrine (bande 2,1) qui ancre le cytosquelette membranaire à la membrane,
à l'hémoglobine,
à plusieurs enzymes glycolytiques.
Band 3 joue un rôle essentiel dans les échanges O2/CO2 au niveau de l'érythrocyte par son mécanisme échangeur d'ions et son action sur la désoxyhémoglobine (interactions avec l'hémoglobine).
Liaisons dans les complexes
membranaires
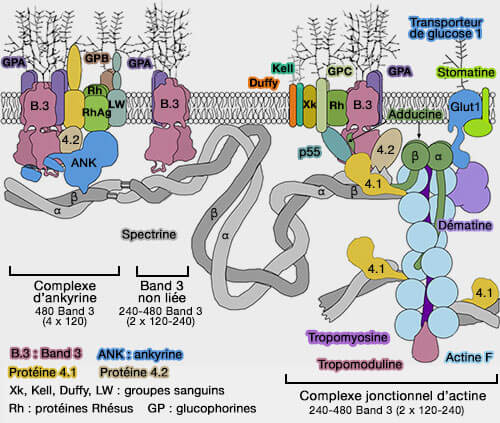
Band 3 entre en interaction avec les complexes qui relient le cytosquelette d'actine à la membrane érythrocytaire par deux ponts différents :
les complexes d'ankyrine,
les complexes jonctionnels d'actine.
Liaison Band 3/Ankyrine
(Figure : vetopsy.fr d'après Grey et coll)
Ces complexes, qui ont un rôle fondamental dans la forme des érythrocytes et la régulation de leurs propriétés mécaniques, sont formés de nombreuses protéines : leur position relative n'est pas connue avec précision et leur proportion est variable, ce qui est logique vu la variabilité des contraintes.
La liaison de l'ankyrine à la spectrine, elle-même liée, par son domaine ABD (Actin Binding Domain) N-terminal, composé de 2 domaines CH (Calponin Homology), à l'actine, permet un premier pont entre la membrane et le cytosquelette.
une boucle comprenant les résidus 175-185 constitue le site d'ancrage,
une boucle adjacente, comprenant des résidus 63-73, contribue aussi à cette liaison.
Cette liaison est réalisée par de nombreux ponts salins et des contacts hydrophobes entre les deux protéines :
Lys69 de Band 3 à Glu645 de l'ankyrine,
Glu72 à Lys611,
Asp183 à Asn601 et Gln634.
D'autres sont moins essentiels : Lys160/His579, Arg155/Asn568, Glu151/Lys535, Thr126/Thr500 et Gln124/Asp 649.
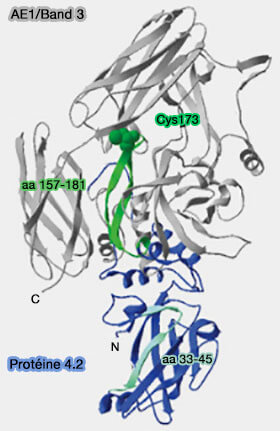
Liaison à la protéine 4.2
La protéine 4.2 (Band 4.2) est une protéine de liaison à l'ATP qui se lie à la fois à Band 3 et à l'ankyrine pour renforcer le lien membrane/cytosquelette.
Elle est myristoylée sur Gly2 et palmitoylée sur Cys173, ce qui permet son ancrage membranaire.
1. La région N-terminale de la protéine 4.2 (résidus 1-238) semble impliquée dans la liaison avec Band 3, mais les études diffèrent sur la localisation exacte du site de liaison.
3. En outre, la protéine 4.2 se lie au complexe jonctionnel d'actine (cf. plus bas).
Complexes jonctionnels d'actine
1. Le squelette de la membrane des globules rouges forme un réseau à peu près hexagonal composé principalement de courts filaments d'actine-F réticulés par des hétérotétramères de spectrine α2/β2 avec l'aide de la protéine 4.1 (Band 4.1 ou EPB41).
Complexes ankyrine et jonctionnel d'actine
(Figure : vetopsy.fr d'après Korsgren et coll)
L'actine et la protéine 4.1 se lient au domaine de liaison à l'actine (domaine ABD formé de 2 domaines CH) situé à l'extrémité N-terminale de la chaîne β de la spectrine.
L'extrémité carboxy-terminale de l'α-spectrine contient un domaine EF (main EF), ressemblant à la calmoduline, avec des EF-mains dépendantes du calcium et indépendantes du calcium dont le rôle n'est pas bien établi.
Le complexe jonctionnel est formé par :
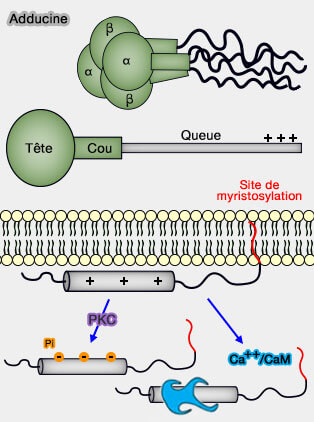
un complexe cytosquelettique formé par la spectrine, l'actine et la protéine 4.1, mais aussi l'adducine, la dématine, la tropomyosine, la tropomoduline et le p55 ;
La protéine 4.2 est située près de la protéine 4.1 aux jonctions spectrine/actine/Band 3, ce qui pourrait fournir un deuxième point de fixation du cytosquelette avec la membrane, en plus de celui de l'ankyrine.
La désoxyHb ne se lie qu'aux résidus 12-23 de Band 3.
Extrémité N-terminale de Band 3
(Figure : vetopsy.fr d'après Shu et coll)
Le site de liaison de Hb sur la bande 3 se trouve à proximité de sites de liaison pour des enzymes glycolytiques, la bande 4.1 et l'ankyrine, ce qui suggère que de multiples propriétés érythrocytaires pourraient être régulées par l'état d'oxygénation de la cellule.
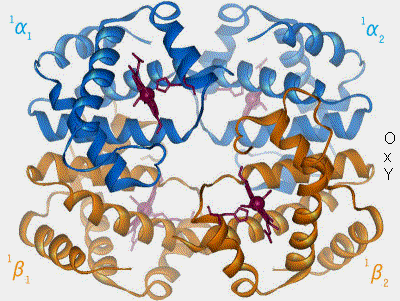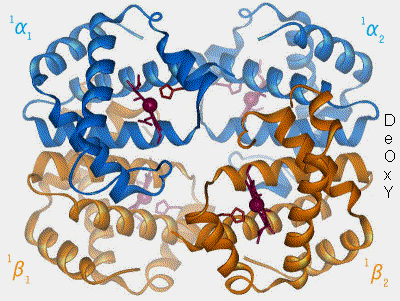
1. La désoxyhémoglobine, forme désoxygénée, possède une conformation dite T ou tendue. Elle possède une faible affinité pour l'oxygène (O2), i.e. elle a tendance à le libérer lorsque le sang circule à travers les tissus.
La forme T est favorisée quand le pH est faible (acide), la concentration en dioxyde de carbone (CO2) forte, un taux élevé en 2,3-bisphosphoglycérate (2,3-BPG) qui stabilise cette forme d'Hb, i.e. favorise la libération d'O2 de la forme R.
Ce segment anionique hautement flexible s'insère parfaitement dans la cavité centrale cationique de la forme T, exploitant la symétrie moléculaire et la polarité positive en formant des ponts salins avec des résidus de lysine et d'histidine dans les sous-unités β de l'hémoglobine (composés à liaison phosphate riche en énergie).
CO2 se lie plus facilement à la désoxyhémoglobine, ce qui facilite son élimination de l'organisme (effet Haldane).
La baisse d'affinité de l'hémoglobine pour l'oxygène en présence de CO2 et de pH acide est appelée effet Bohr.
Passage de la forme T à la forme R de l'hémoglobine
(Animation : BerserkerBen)
2. L'oxyhémoglobine, à forme R ou relâchée, a exactement les caractéristiques inverses, i.e. forte affinité pour l'oxygène (O2), ce qui favorise la fixation de l'oxygène lorsque le sang circule au niveau des alvéoles pulmonaires.
Lors de la liaison à O2, la cavité centrale est liée au 2,3-BPG, i.e. son volume diminue fortement, ce qui explique le manque d'affinité de l'oxyhémoglobine pour Band 3.
La consommation de glucose érythrocytaire se produit principalement par l'intermédiaire de la glycolyse dans les cellules désoxygénées.
Lors de l'exposition à O2, une très grande partie du glucose cellulaire est métabolisée par la voie des pentoses phosphates ou voie de Warburg-Dickens-Horecker qui produit essentiellement, pour notre sujet, du NADPH (Nicotinamide adenine dinucleotide phosphate, réducteur) et un proton (H+).
Ainsi, en activant la voie du pentose phosphate lors de l'oxygénation des érythrocytes, la cellule est assurée d'une quantité suffisante de NADPH pour la réduction du glutathion, puissant antioxydant, et le maintien de l'hémoglobine en oxyhémoglobine.
Ces enzymes glycolytiques sont en compétition directe avec le désoxyHb pour l'extrémité NH2 de la bande 3, les enzymes sont déplacées durant la désoxygénation des érythrocytes.
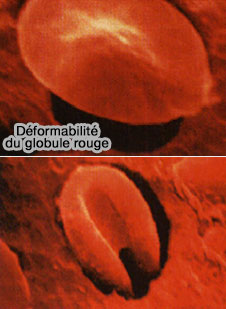
Déformabilité de l'érythrocyte
(Figure : rrmechatronics)
2. Ce mécanisme peut aussi expliquer pourquoi l'état d'oxygénation des globules rouges peut moduler les propriétés mécaniques de la membrane (déformabilité des érythrocytes).
Pendant leur vie de 120 jours, les globules rouges sont pressurés à travers les capillaires ou les sinusoïdes qui mesurent moins de la moitié de leur diamètre.
Les 2 boucles de liaison à l'ankyrine sur cdb3 sont situées à moins de 15 Å du site de liaison de désoxyHb.
L'ankyrine devant être sphérique (100 Å environ) comme la désoxyHb (55 Å environ), il paraît plus que probable que la désoxyHb liée à la Band 3 déplace l'ankyrine.
Les périodes de désoxygénation conduiraient donc à la rupture des ponts entre cytosquelette et membrane, provoquant une déformabilité accrue des globules rouges désoxygénés pour qu'ils atteignent plus rapidement les poumons.




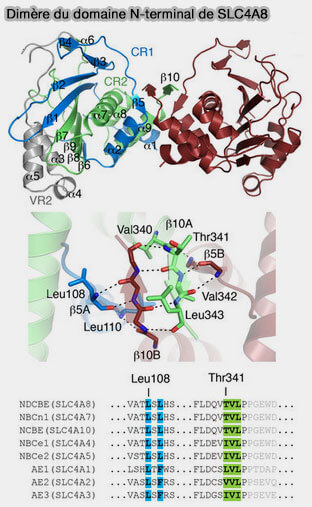
 dimérisation des domaines membranaires).
dimérisation des domaines membranaires).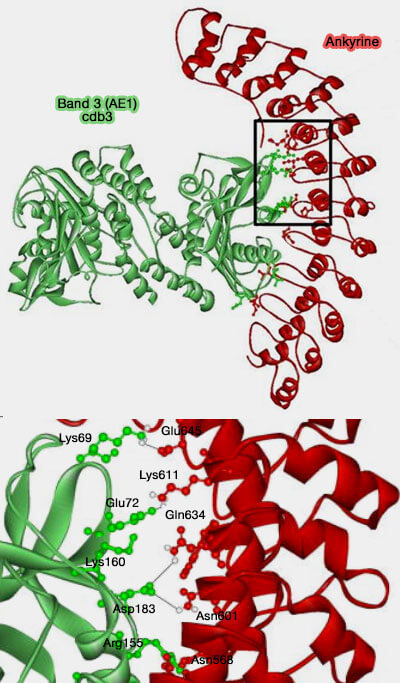


{kind=link}