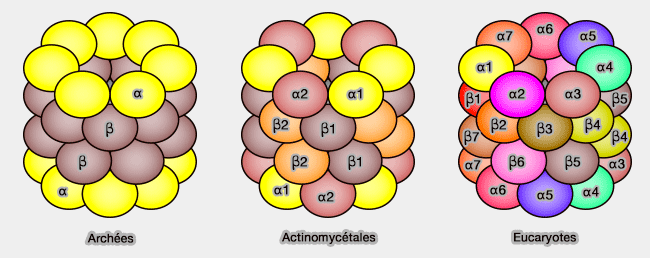
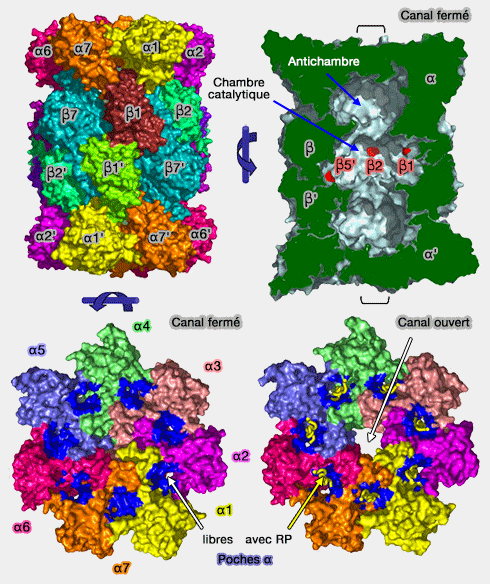
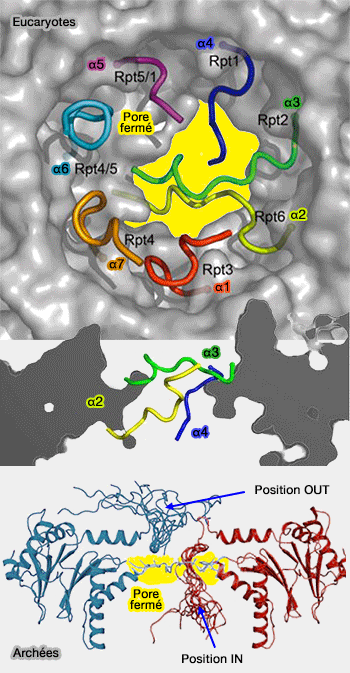
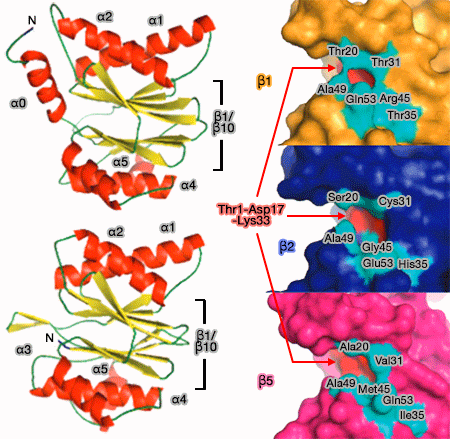
Coeur catalytique 20S (CP) du protéasome
(Figure : vetopsy.fr d'après Finley)7 sous-unités α extérieures, sites de liaison des complexes régulateurs, qui forment une antichambre,
7 sous-unités β intérieures qui forment, avec le deuxième anneau β, la chambre catalytique du complexe protéasique.
Chez les Archées, les sept sous-unités α et β sont semblables alors qu'elles sont toutes différentes chez les Eucaryotes (Structure of the Proteasome 2012).
1. Le pore d'entrée des antichambres est occlus par les extrémités N-terminales des sous-unités α et empêche le substrat de pénétrer dans l'antichambre.
Cette antichambre mesure 40 Å sur 50 Å pour 59 nm3 chez T. acidophilum.
Chez les archées, dont toutes les sous-unités sont identiques, les 12 résidus N-terminaux très flexibles de chaque sous-unité sont situés dans le pore et s'étendent à travers l'anneau (aussi bien dans l'antichambre qu'à l'extérieur) pour empêcher l'accès aux protéines (Dynamic Regulation of Archaeal Proteasome Gate Opening As Studied by TROSY NMR 2010).
Fermeture du pore
(Figure : vetopsy.fr d'après Finley et Religa)
Chez les eucaryotes, comme toutes les sous-unités α sont différentes, ce sont les extrémités N-terminales des sous-unités α2, α3 et α4 qui interviennent dans ce processus. Les différences observées dans les Rpt de la base du complexe 19S et les interactions de leur extrémités C-terminales dans les poches entre les sous-unités α, pourraient expliquer leur rôle spécifique (cf. translocation détaillée du substrat).
2. Lors de son ouverture, après contact avec un substrat, le pore d'entrée ne fait que 13 Å de large.
La chambre catalytique est formée par les sous-unités β.
La chambre catalytique fait 53 Å de large pour 84 nm3 de T. acidophilum.
Le substrat doit passer par le pore étroit de l'anneau α (13 Å de large lors de son ouverture), ce qui protège, par ailleurs, les autres protéines cellulaires de toute dégradation intempestives.
D'une part, l'activité protéolytique du protéasome est cloisonnée à l'intérieur de son enveloppe protéique, un peu à l'instar d'un compartiment cellulaire comme celui des mitochondries.
Le propeptide est clivé pour révéler un résidu thréonine à son extrémité N-terminale (Thr1).
Au cours de la protéolyse, le polypeptide se lie au groupe hydroxyle N-terminal de la thréonine, probablement via des liaisons hydrogène. La triade catalytique (nucléophile, acide, base) est formée de Thr1, Asp17 (au lieu de Glu17 des bactéries) et Lys33.
Activité protéolytique du coeur catalytique du protéasome
(Figure : vetopsy.fr d'après Marques)
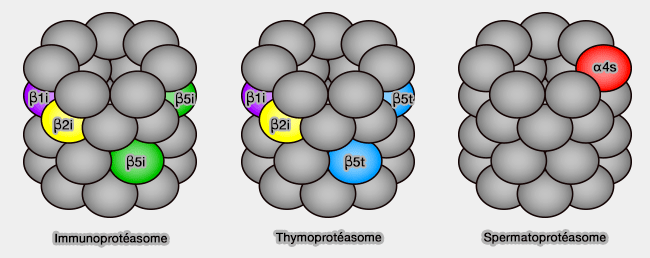
Trois sites actifs sur chaque anneau, β1, β2 et β5, peuvent cliver à peu près toutes les protéines après des résidus :
hydrophobes (β5 à activité chymotripsique : protéase à sérine qui clive les protéines après les acides aminés aromatiques ou hydrophobes tels que tyrosine, phénylalanine ou encore tryptophane). C'est la seule activité qui existe chez les Procaryotes.
Les caractéristiques des poches à protéolyse sont largement conditionnées par le résidu 45 : Arg (basique), Gly (apolaire) et Met (apolaire), respectivement pour β1, β2 et β5.
La plupart des inhibiteurs protéasomaux se lient à β5, agissant donc sur l'activité chymotrypsine, sans modifier les deux autres.