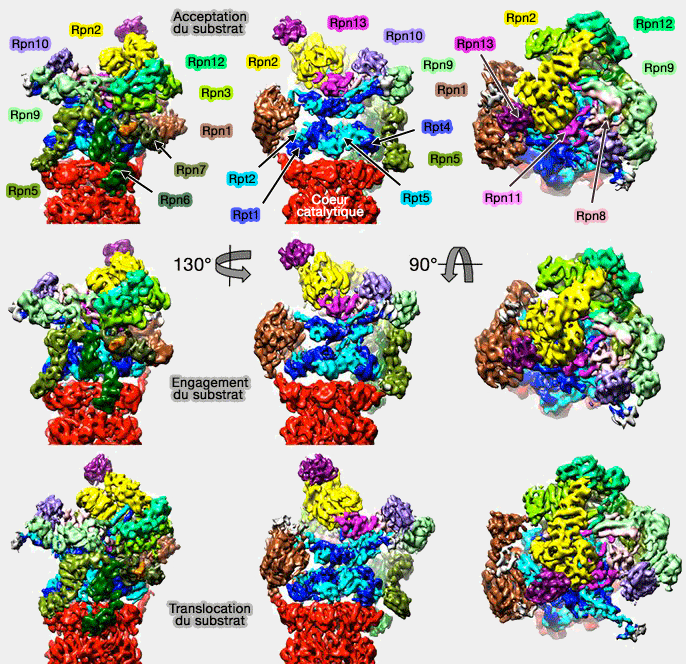
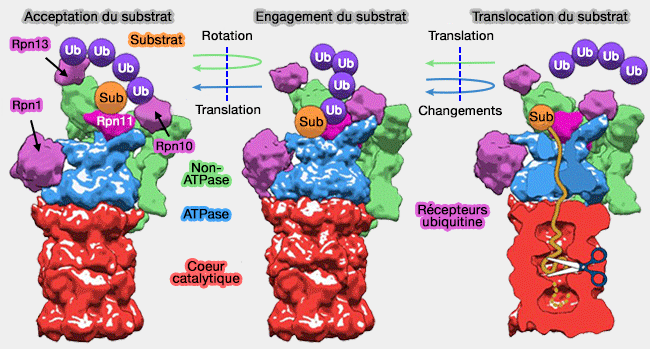
Une fois l'acceptation du substrat et son engagement effectué, le protéasome déplace la chaîne polypeptidique du substrat vers la chambre catalytique.
Le déplacement de la base (OB et ATPase), qui suit le déplacement du " couvercle " (lid) par la liaison avec la chaîne polyubiquitinée du substrat, se poursuit par un pivotement de telle sorte que tous les anneaux deviennent alignés de façon coaxiale pour former un canal continu.
Vous pouvez aussi voir deux films tirés de cet article : sur les changements de conformation vus de côté et vus du haut du protéasome.
Acceptation, engagement et translocation du substrat
(Figure : vetopsy.fr d'après Unverdorben)
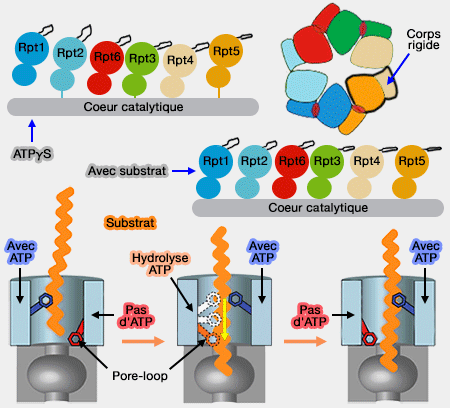
Translocation simplifiée du substrat
Anneau N
L'anneau OB ou N, structure rigide, est en forme d'entonnoir avec un anneau central comportant un anneau proximal (13 Å) bien plus étroit que l'anneau distal.
Fonctionnement du moteur ATPase dans la translocation
(Figure : vetopsy.fr d'après Martin et Bhattacharyya)
Le dépliage du substrat est probablement le résultat d'une translocation forcée à travers cet anneau.
La boucle Ar-Φ (Aromatic-aliphatic ou pore-1 loop) - Y/F-I/L/V-G -, leur permet de se lier à la protéine, de la déplier et de la transloquer au CP : ces boucles font saillies dans le pore axial de l'anneau AAA+
La pore-2 loop, situé à côté de la boucle Ar-Φ, semble aussi jouer impliquer dans ces processus. Il semble qu'elle entre aussi en interaction avec le coeur catalytique.
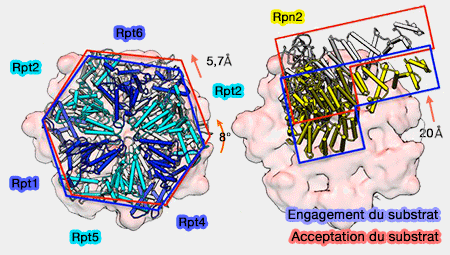
Changements lors de la translocation du substrat
(Figure : vetopsy.fr d'après Unverdorben)
L'anneau ATPase se positionne parallèlement au sommet de l'anneau α du coeur catalytique : translaté (≅6Å), en rotation latérale (≅8°), et moins incliné (≅-3°), ce qui aligne ces deux anneaux.
Il pourrait jouer un rôle dans la liaison base/coeur.
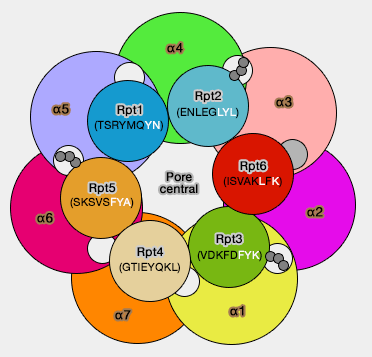
En effet, c'est le seul qui montre une nette spécificité pour une poche particulière (α2/α3) par rapport aux autres Rpt, et serait aussi le point de départ de l'assemblage par les protéines chaperonnes, ce qui éviterait par exemple, aux autres de se fixer à un mauvais endroit.
Conséquences
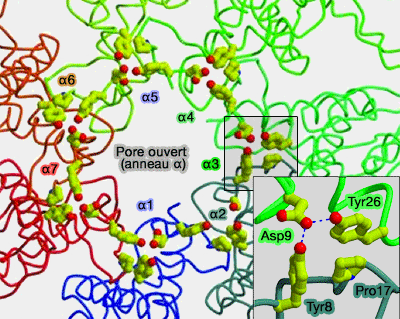
Cette liaison provoque une rotation des sous-unités et l'ouverture du pore.
Ouverture du pore (PAN)
(Figure : vetopsy.fr d'après Fôrster)
Cela se traduit par la formation d'un complexe de quatre résidus hautement conservés, Tyr8, Asp9, Pro17 et Tyr26, entre chacune des sous-unités adjacentes (A gated channel into proteasome core particle 2000).
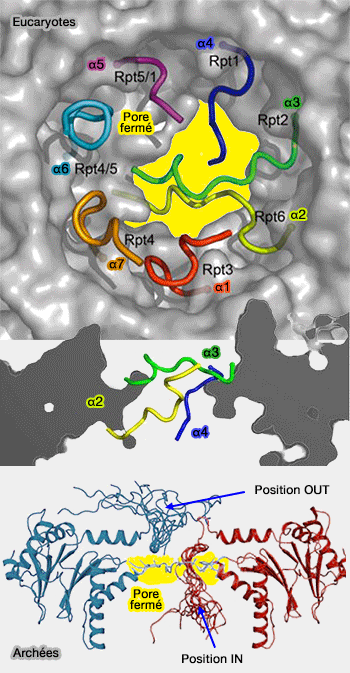
2. Chez les eucaryotes, le mouvement de basculement des sous-unités α ne nécessite pas forcément la boucle d'activation, ni l'interaction avec Pro17, mais utilise plutôt les interactions de leur résidu C-terminal.
La liaison du nucléotide modifie la rotation entre les grands et les petits domaines AAA+ des Rpt, et expliquent le mécanisme permettant de relier le cycle ATPasique aux changements conformationnels dans l'anneau hexamérique des Rpt, à l'alignement des anneaux et, enfin à la translocation du substrat.
En outre, les réarrangements conformationnels de l'anneau ne semblent pas non plus être indispensables. Dans ClpX, l'anneau peut rester fermé sans que cela affecte beaucoup la dégradation de la protéine.
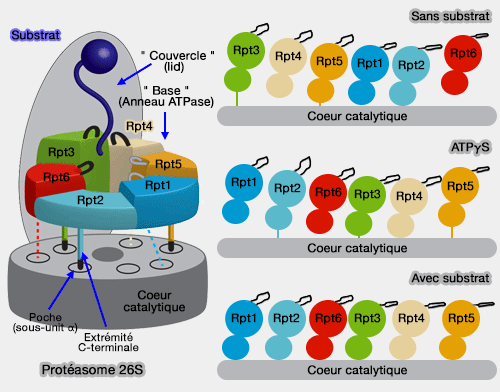
Liaison de l'anneau ATPase avec le coeur catalytique (simplifié)
(Figure : vetopsy.fr d'après Estin et Bhattacharyya)
Il semble que ce stade corresponde au stade où un substrat s'engage dans le pore central, stade long (" dwell "), environ 90% du temps, au cours duquel l'ADP est relarguée et les Rpt rechargés en ATP.
Le reste (10% du temps) est une phase de bouffée (" burst "), au cours de laquelle le substrat avance d'une certaine longueur à travers le pore, grâce aux pore-loop : cette progression est la conséquence des changements conformationnels coordonnés des sous-unités autour de l'anneau par l'hydrolyse rapide et progressive des différentes Rpt (High Degree of Coordination and Division of Labor among Subunits in a Homomeric Ring ATPase 2012).
Puis le cycle recommence jusqu'à la translocation complexe du substrat.
Quoi qu'il en soit, il semble que les sous-unités ATPases agissent de manière coordonnée pour hydrolyser l'ATP et ainsi propulser des substrats à travers le canal de translocation.
Acceptation, engagement et translocation du substrat
(Figure : vetopsy.fr d'après Unverdorben)
Positionnement de l'anneau OB
L'anneau OB ou N est plus translaté de 17,5Å que l'anneau ATPase : Rpn1 se déplace de manière identique.
Les autres modifications sont minimes.
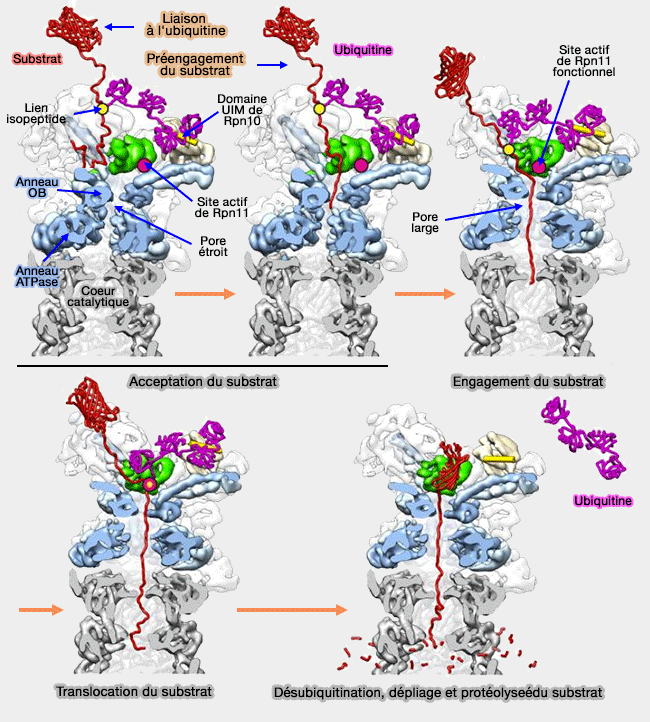
Ces processus permettent la translocation du substrat, sa désubiquitination et le recyclage de l'ubiquitine, et sa protéolyse par le coeur catalytique en oligopeptides de 7 à 8 résidus.
Fonctionnement complet du protéasome
(Figure : vetopsy.fr d'après Lander)