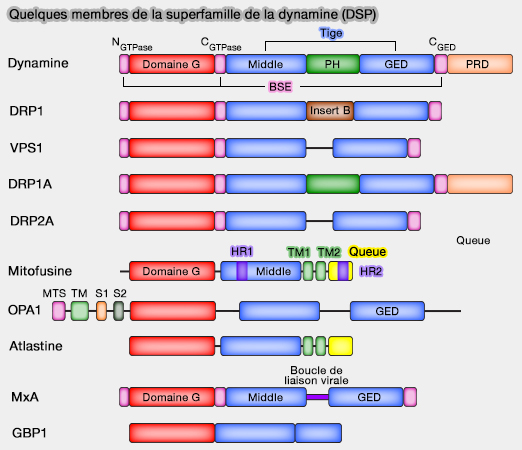
Quelques membres de la superfamille de la dynamine
(Figure : vetopsy.fr)
Domaine G
Le domaine G contient des insertions qui intensifient le repliement classique et leur confèrent des propriétés biochimiques et biophysiques spécifiques.
la dimérisation se produit entre les bicouches membranaires apposées.
Leur interface beaucoup plus réduite pourrait expliquer la faible affinité pour les nucléotides guanidiques et un taux d'hydrolyse GTPasique beaucoup plus faible.
Les tiges, par ces interactions membranaires, contrôlent par un mécanisme inhibiteur l'auto-assemblage.
Ces ancrages obstruent stériquement l'accès à l'interface 3 et empêche l'auto-assemblage prématuré de ces DSP en l'absence d'interactions spécifiques de la membrane cible.
L'association stable avec la membrane expose l'interface 3.
DSP de fusion
1. Les DSP de fusion sont des monomères qui se dimérisent de manière réversible via leurs domaine G et leurs tiges par une coopération active des nucléotides pour former des réseaux hélicoïdaux ou plats.
La base de HB2 est connectée à des éléments qui s'intègrent à la bicouche membranaire (comme les segments transmembranaires - TM - de Mfn1/2 et la palette de BDLP)
Mitofusine1 (Mfn1) et BDLP
(Figure : vetopsy.fr d'après Qi et coll)
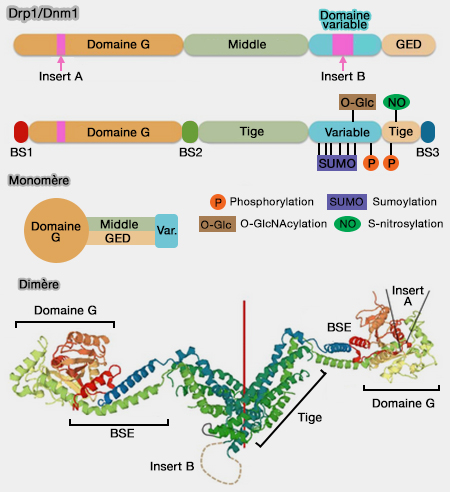
Le BSE subit un réarrangement conformationnel par rapport au domaine G au cours des cycles d'hydrolyse coopérative du GTP (modèles de fission de la dynamine).
Les DSP dits " anciens ", i.e. phylogénétiquement, comportent des inserts polypeptidiques intrinsèquement désordonnés, appelés aussi domaines variables, qui jouent un rôle dans régulation de leur fonction comme dans :
Monomère de dynamine
(Figure : vetopsy.fr d'après Antonny et coll)
Vps1p de la levure se lie aux lipides et aux partenaires de liaison à domaine SH3 pour remodeler la membrane.
Ces inserts des " anciens " DRP sont les domaines précurseurs fonctionnels de la dynamine, DSP " moderne " :
le domaine PH, retrouvé dans la pleckstrine, protéine des plaquettes, qui se lie au PI(4,5)P2 ou PIP2 membranaire, et régule aussi, par ses boucles hydrophobes, l'autoassemblage et la fission, comme le fait l'insert B de Drp1.
Structures des différentes formes d'Opa1
(Figure : vetopsy.fr)
2. OPA1, comme les protéines de fission (dynamine et Drp1) contient un domaine " middle " et un domaine GED connectés par une région polypeptidique intrinsèquement désordonnée qui rappelle l'insert B Drp1.
L'OPA1, dans sa version longue (L-OPA1), ancré dans le domaine transmembranaire et reliant les deux surfaces mitochondriales internes, permettrait la fusion par des interactions membranaires secondaires régulées par le domaine variable.
L'ancre transmembranaire subit une excision protéolytique régulée pour libérer une forme soluble plus courte (S-OPA1) qui peut fonctionner dans la fission membranaire ( formes d'OPA1).
En effet, l'activité de l'OPA1 a été impliquée à la fois dans la fusion de la membrane interne mitochondriale et dans la fission





 modèles de fission et partenaires), ce chapitre traite des similitudes et des différences des structures des autres DSP par rapport à cette GTPase.
modèles de fission et partenaires), ce chapitre traite des similitudes et des différences des structures des autres DSP par rapport à cette GTPase.