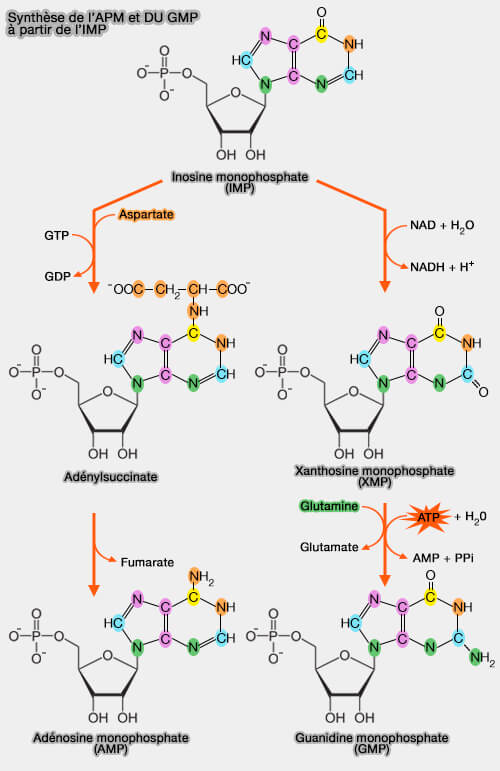
Les ribonucléotides puriques AMP et GMP sont formés à partir de l’inosine monophosphate (IMP), nucléotide précurseur commun dont les voies de transformation permettent d’équilibrer la biosynthèse des purines.
Ainsi, l’IMP constitue un point de branchement majeur du métabolisme des purines, permettant la formation de l’AMP et du GMP par deux voies enzymatiques distinctes impliquant respectivement l’aspartate et la glutamine.
Formation de novo de l'AMP à partir de l'IMP
Vue d'ensemble
La formation de novo de l'AMP constitue l'étape suivante de la formation de novo de l'IMP, nucléoside princeps dans cette voie.
a. C'est l'aspartate qui cède l'atome d'azote par un mécanisme semblable à celui permettant l'introduction de l'atome d'azote (N1) lors de la synthèse de l'IMP (réaction 7 et 8), qui formera l’azote N6 de l’adénine.
b. Cette réaction est énergétiquement coûteuse, l’énergie étant fournie par l’hydrolyse du GTP, dont l’utilisation contribue à la régulation croisée de la biosynthèse des nucléotides puriques, la synthèse de l’AMP dépendant ainsi du niveau intracellulaire de GMP/GTP.
L'adénylosuccinate synthétase subit une rétro-inhibition par l'AMP, mécanisme de régulation allostérique classique dans la biosynthèse des purines.
Nucléoside-diphosphate kinase
(Figure : vetopsy.fr)
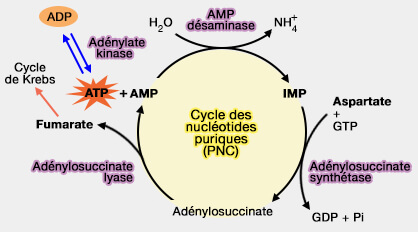
le maintien d'un équilibre dynamique entre les nucléotides AMP, IMP et les acides aminés, notamment l’aspartate et le fumarate.
a. Ce cycle comprend, en aval des deux étapes précédentes, la formation de l'IMP à partir de l'AMP catalysée par l’AMP désaminase(EC 3.5.4.6).
b. Cette réaction est très active dans le muscle, notamment lors d’un effort, pour éviter l’accumulation d’AMP.
Ce cycle, qui permet de régénérer l’AMP sans perte nette, tout en produisant du fumarate, un métabolite intermédiaire du cycle de Krebs (étape 8), joue un rôle central à l’interface entre le métabolisme des nucléotides puriques, des acides aminés et des voies énergétiques.
Formation du GMP à partir de l'IMP
La formation de novo du GMP constitue l'étape suivante de la formation de novo de l'IMP, nucléotide princeps dans cette voie.
La synthèse se fait en deux étapes.
1. L'IMP est oxydé au niveau du carbone 2 de la base purique pour former la xanthosine-monophosphate (XMP ou acide xanthylique), nucléotide à base de xanthine.
a. La réaction est catalysée par l'IMP déshydrogénase (inosine-5'-monophosphate déshydrogénase), EC 1.1.1.205, enzyme clé et souvent limitante dans la biosynthèse des nucléotides guanyliques ( régulation).
Le coenzyme $\ce{NAD+}$ accepte les électrons issus de l’oxydation, formant du NADH et un proton $\ce{H+}$,
2. Dans un second temps, on assiste à une amidation du carbone 2, i.e. remplacement du $\ce{=O}$ du carbone 2 par $\ce{-NH2}$, l'azote provenant du groupe amide ($\ce{-C(=O)NH2}$) de la glutamine.
a. Cette amidation n'est pas une amidation classique comme celle entre un acide carbonique et une amine mais une amidation qu'on peut qualifier de fonctionnelle.
$\ce{XMP + ATP + L-glutamine + H2O}$
$\leftrightharpoons$ $\ce{GMP + AMPP + PPi + L-glutamate}$
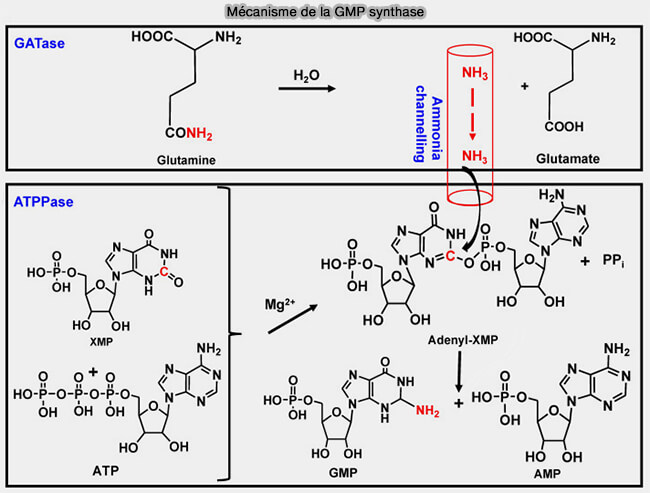
Mécanisme de la GMP synthase
(Figure : vetopsy.fr d'après Ballut et col)
activation du groupe cétone($\ce{-C=O}$) en C2 de la xanthine, faiblement électrophile, par l'ATP, pour former un XMP-AMPylé, fortement électrophile, à l'aide classique de Mg++,
hydrolyse de la glutamine en glutamate et NH3 qui est acheminé à travers un canal protéique interne jusqu’au site d’activation du XMP (mécanisme de canalisation de substrat),
d. La GMP synthétase subit une rétro-inhibition par le GMP, mécanisme de régulation allostérique classique dans la biosynthèse des purines.
L'utilisation du GTP pour la synthèse d'AMP, et de l'ATP pour la synthèse de GMP permet de comprendre pourquoi les concentrations des deux ribonucléotides puriques sont comparables.
L'accumulation de GTP accélère la synthèse d'AMP à partir de l'IMP au détriment de la synthèse de GMP.
À l'inverse, l'accumulation d'ATP conduit à la synthèse accélérée du GMP au détriment de la synthèse d'AMP.
Transformation des nucléotides puriques obtenus par biosynthèse de novo
Les nucléosides monophosphates puriques formés lors de la biosynthèse de novo, i.e. l’AMP et le GMP, peuvent subir des phosphorylations successives conduisant à la formation des nucléosides diphosphates et triphosphates correspondants.
1. D'une part, les nucléosides mono-, di- et triphosphates puriques peuvent également être interconvertis par phosphorylation et déphosphorylation enzymatiques, permettant le passage réversible entre les différentes formes phosphorylées (biosynthèse des NDP et des NTP).
Remarque : les nucléosides diphosphates peuvent ensuite être réduits en désoxyribonucléosides diphosphates par la ribonucléotide réductase, étape essentielle pour la synthèse de l’ADN (biosynthèse des désoxyribonucléotides).





 régulation).
régulation).