


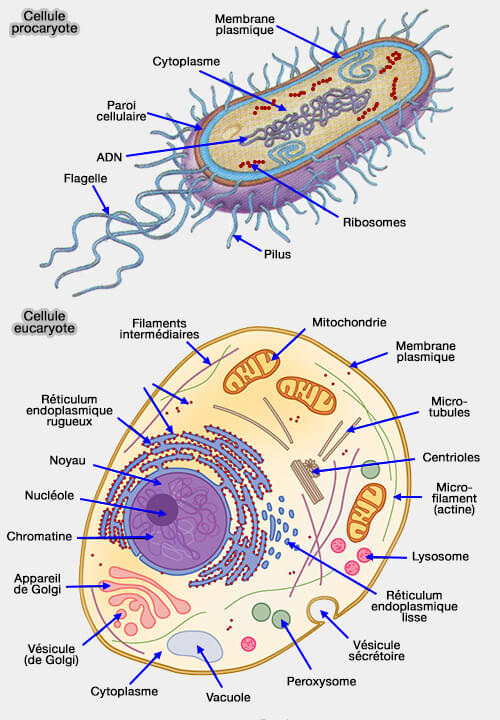
Biologie cellulaire
Théorie endosymbiotique : origine des organites
- Biologie cellulaire et moléculaire
- Constituants de la cellule
- Matrice extracellulaire
- Reproduction cellulaire
- Biochimie
- Transport membranaire
- Moteurs moléculaires
- Voies de signalisation

La théorie endosymbiotique (symbiogénose ou symbiogenèse) est une théorie évolutionniste qui soutient que les organites des eucaryotes sont issus de l'endosymbiose de procaryotes comme des bactéries ou des archées.
- L'endosymbiose, du grec endon " dans ", syn " ensemble " et biosis " vivant ", est la coopération mutuellement bénéfique entre deux organismes vivants, i.e. une forme de symbiose, où l'un est contenu par l'autre.
- L'organisme interne est appelé endosymbiote (endosymbionte ou endobionte).
Théorie endosymbiotique et organites
Ces organites seraient :
- le réticulum endoplasmique et les membranes nucléaires ;
- les mitochondries qui dériveraient des rickettsiales, une classe de l'ordre des alphaproteobacteria appartenant au phylum des Proteobacteria, phylum majeur des bactéries à gram négatif ;
- les plastes (ou plastides), présent dans les cellules eucaryotes chlorophylliennes (algues et plantes) comme les chloroplastes qui seraient apparentés aux cyanobactéries (algues bleues) filamenteuses fixant l'azote.
Cette théorie a été énoncée en 1905 par le botaniste russe Konstantin Mereschkowski (1855-1921), puis étayée en 1967 par la microbiologiste américaine Lynn Margulis (1938-2011) dans son article : On the Origin of Mitosing Cells.
- Lynn Margulis fait de cette coopération entre organismes un des moteurs de l'évolution en réaction au néo-darwinisme, vision pour elle négative dans laquelle la lutte pour la survie est une guerre incessante.
- Elle pensait que les néo-darwiniens : « vautrés dans leur interprétation de Darwin du point de vue de la zoologie, du capitalisme, de la compétition, des coûts-bénéfices - ont complètement faussé sa pensée. Le néo-darwinisme, qui insiste sur l'accumulation lente de mutations par la sélection naturelle au niveau du gène, est une théorie de trouillards. »
-
Ses prises de position postérieures sur d'autres sujets se sont révélées plus que contestables (
 controverses).
controverses).

Pour tout savoir sur cette théorie lire : Endosymbiotic theories for eukaryote origin (2015), Endosymbiosis and Eukaryotic Cell Evolution (2015), Endosymbiotic origin and differential loss of eukaryotic genes (2015) et Endosymbiotic theory for organelle origins (2014).
Preuves étayant la théorie
Dans cette partie, on ne s'intéressera qu'aux mitochondries, sachant que les chloroplastes partagent avec elles de nombreuses similitudes.
(Figure : vetopsy.fr d'après Archibald et coll)
1. Les mitochondries contiennent leur propre l'ADN circulaire à double brin, l'ADN mitochondrial (ADNmt) , plus proche d’un génome procaryote et différent et beaucoup plus réduit que l'ADN nucléaire (génome mitochondrial).
Ce génome est très proche de celui de Rickettsia prowazekii, agent du typhus exanthématique (The genome sequence of Rickettsia prowazekii and the origin of mitochondria 1998).
2. Ces organites sont entourés de deux membranes.
- La membrane externe (OMM) est presque identique aux membranes des cellules eucaryotes.
- La membrane interne (IMM) est proche de celle des bactéries à gram négatif, en particulier par la présence d'un lipide particulier, la carbolipine.
3. Les nouvelles mitochondries se forment uniquement par fission, cette division étant indépendante du cycle nucléaire (fusion et fission des mitochondries).
4. Les ribosomes des mitochondries (mitoribosomes) sont proches de ceux des bactéries, en particulier par leur sensibilité aux antibiotiques.
Comment s'est effectuée cette endosymbiose ?
Modèles
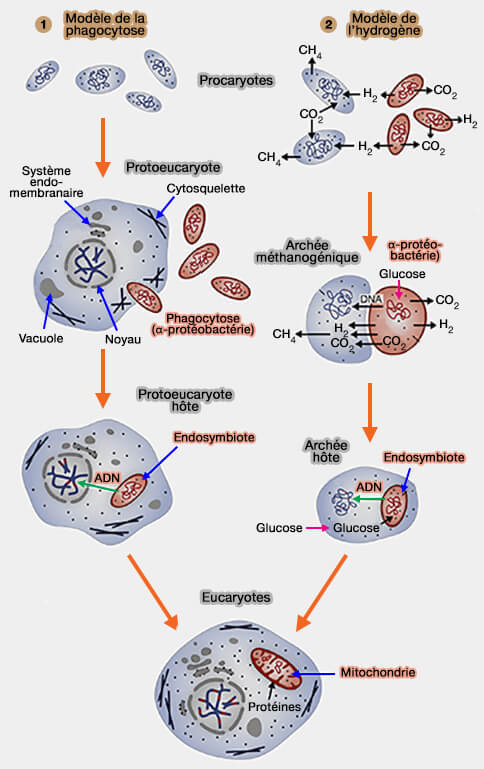
De nombreux modèles ont été élaborés dont deux principaux pour expliquer l'origine des mitochondries (Endosymbiosis and Eukaryotic Cell Evolution 2015).
1. Le premier modèle suppose une phagocytose incomplète par une cellule protoeukaryote d'une bactérie comme l'alphaprotéobacterie Rickettsia prowazekii aérobie citée plus haut.
Ce modèle implique que les archées, i.e. autotrophes et anaérobies, ont grossi et l'invagination de leur membrane a formé :
- le réticulum endoplasmique,
- le noyau.
L'endosymbiote, hétérotrophe, mais aérobie, apporte cette dernière particularité à la cellule protoeucaryote hôte devenue maintenant cellule eucaryote.
- L'endosymbiote perd des gènes, en transfère au noyau : son système d'importation de protéines évolue.
- L'endosymbiote devient organite.
2.. Le deuxième modèle ou modèle de l'" hydrogène " implique la symbiose métabolique entre (1 de la figure ci-dessous).
- une archée qui produit du méthane (qui dépend de H2) et
- une alphaprotéobactérie capable de respirer en présence d'O2, ou effectuer des fermentations produisant H2 dans des conditions anaérobies. (The hydrogen hypothesis for the first eukaryote 1998).
Dans ce modèle, la complexité des cellules eucaryotes apparaît après l'endosymbiose.

Toute la force de ce modèle s'exprime dans le fait que l'hôte a réellement besoin de son endosymbionte mitochondrial. La suite de ce chapitre est tiré de Endosymbiotic theories for eukaryote origin (2015).
Modèle de l'" hydrogène "
Origine des mitochondries
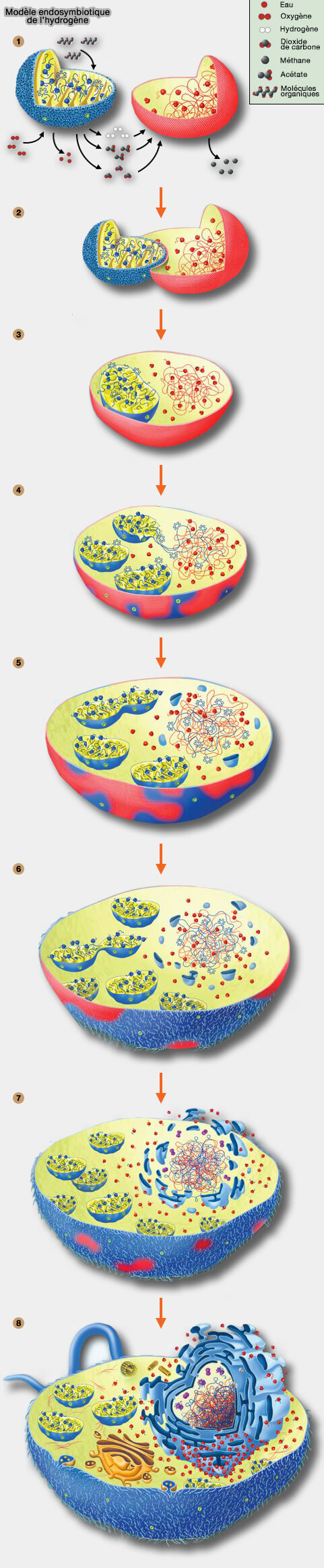
La syntrophie anaérobie, i.e. du grec syn, ensemble, et trophe, nourriture, phénomène qu'une espèce vit des produits d'une autre espèce, ici, transfert de H2 serait donc le contexte métabolique de l'association hôte-symbiote, conduisant à des hôtes qui ont tendance à interagir étroitement (2 de la figure ci-dessous) avec leurs symbiotes (Archaeal phylogenomics provides evidence in support of a methanogenic origin of the Archaea and a thaumarchaeal origin for the eukaryotes 2010).
On retrouve des associations symbiotiques similaires entre les méthanogènes, i.e. archées qui produisent du méthane sous forme de sous-produit métabolique dans des conditions anaérobies, dans les hydrogénosomes du cytosol de ciliés anaérobies (A new polymorphic methanogen, closely related to Methanucorpuscuhm parvum, living in stable symbiosis within the anaerobic ciliate Trimyema sp. 1993).
La nature H2-dépendante de l'hôte conduit à une situation curieuse (3 de la figure ci-dessous).
- Afin de générer de l'H2 pour l'hôte, l'endosymbiote nécessite des composés organiques réduits (substrats organiques fermentescibles), mais l'hôte est un autotrophe strict, i.e. sans prélèvement de molécules organiques dans le milieu.
- Il ne peut pas en fournir plus que ses propres besoins : les autotrophes H2 dépendent du CO2 et de l'eau et n'importent pas de composés organiques réduits.
(Figure : vetopsy.fr d'après Martin et coll)
1. Cette phase de la symbiose est donc instable car le symbiote finira par consommer le cytosol de l'hôte. Pour que la symbiose persiste :
- soit l'hôte doit " inventer " des importateurs de composés organiques,
- soit les gènes préexistants des importateurs de l'endosymbiote doivent être transférés aux chromosomes de l'hôte pour que ces transporteurs soient exprimés sur la membrane de l'hôte.
Dans les haloarchaea, les importateurs bactériens doivent être fonctionnels dans la membrane archéale (Acquisition of 1,000 eubacterial genes physiologically transformed a methanogen at the origin of Haloarchaea 2012).
Ce transfert de gènes pourrait simplement impliquer la lyse occasionnelle d'un endosymbiote, tout comme il se produit dans le transfert de gènes des organites au noyau chez les eucaryotes aujourd'hui (Endosymbiotic gene transfer: organelle genomes forge eukaryotic chromosomes 2004), sauf qu'à ce stade de la symbiose, l'hôte est encore une archée et ne possède pas de noyau (4 de la figure ci-contre).
2. L'expression des importateurs de carbone dans la membrane de l'hôte ne résout cependant pas complètement le problème, car l'hypothèse de l'hydrogène postule que l'hôte était autotrophe, par conséquent son métabolisme du carbone était spécialisé dans les voies anaboliques.
Un bon exemple d'une telle spécialisation enzymatique est la fructose 1,6 bisphosphate aldolase/bisphosphatase caractéristique des archées autotrophes, mais qui n'existe plus chez les eucaryotes (Fructose 1,6-bisphosphate aldolase/phosphatase may be an ancestral gluconeogenic enzyme 2010). On peut en citer d'autres (Carbohydrate Metabolism in Archaea: Current Insights into Unusual Enzymes and Pathways and Their Regulation 2014).
Deux hypothèses sont alors possibles.
- Soit les enzymes du métabolisme anabolique de l'hôte doivent acquérir, une mutation ponctuelle à la fois, les substitutions nécessaires pour réduire le métabolisme du carbone, ce qui prend du temps.
- Soit, un processus bien plus rapide consiste en ce que les gènes du métabolisme hétérotrophique du carbone de l'endosymbiote se tranfèrent dans les chromosomes de l'hôte, les protéines s'exprimant directement dans son cytosol.
Ce transfert est d'une importance capitale pour les deux symbiotes.
- Le flux du carbone est dirigé vers l'endosymbiote, de sorte qu'il peut produire de l'H2 par fermentation pour satisfaire l'hôte.
- Le cytosol de l'hôte devient hétérotrophe, mais seulement si le transfert de la voie glycolytique complète de l'endosymbiote est réussi, i.e. les étapes enzymatiques jusqu'au pyruvate qui conduise au gain d'ATP (bilan énergétique de la glycolyse).
- Ce processus expliquerait directement l'origine bactérienne des enzymes glycolytiques eucaryotes, excepté l'énolase impliquée dans la réaction 9 de la glycolyse (Enolase from Trypanosoma brucei, from the Amitochondriate Protist Mastigamoeba balamuthi, and from the Chloroplast and Cytosol of Euglena gracilis: Pieces in the Evolutionary Puzzle of the Eukaryotic Glycolytic Pathway 2000).
Aucun autre modèle de la théorie endosymbiotique ne permet d'observer que les eucaryotes, dont les ribosomes proviennent d'archées, possèdent une voie glycolytique bactérienne.
L'hypothèse CoRR de John F. Allen (Co-localisation pour la Régulation Redox) postule que les organites ont conservé des génomes de sorte que les organites individuels peuvent avoir leur mot à dire dans l'expression des composants des chaînes de transport d'électrons respiratoire et photosynthétique afin de maintenir l'équilibre redox dans la membrane bioénergétique (The CoRR hypothesis for genes in organelles 2017).
Cela explique la convergence du contenu génétique des mitochondries et des plastes qui ont conservé le codage :
- des gènes impliqués dans leurs chaînes de transport d'électrons respectives,
- des composants des ribosomes nécessaires pour les exprimer dans l'organite.
Cette convergence de la teneur en protéine ribosomique dans les génomes mitochondrial et plastique peut être expliqué par l'assemblage des ribosomes : le processus de la biogenèse des ribosomes nécessite que certaines protéines doivent être co-exprimées dans le même compartiment que leurs ARNr naissants.
3. L'expression des gènes du flux de carbone hétérotrophique dans le compartiment hôte lui confère des molécules carbonées réduites et de l'ATP, ce qui lui permet d'abandonner son autotrophie.
- L'hôte est devenu un hétérotrophe avec des chromosomes chimériques contenant des gènes archéens et bactériens et des ribosomes archéens, et possédant une glycolyse dans le cytosol.
- En outre, le cytosol héberge maintenant un endosymbiote bactérien anaérobie facultatif avec une chaîne respiratoire et des fermentations produisant de l'H2.
Cet endosymbiote peut toujours transférer un génome complet, remplaçant de nombreuses voies archéales par des contreparties bactériennes, et ainsi transformer l'archée de l'intérieur.
Une partie de cette transformation implique la synthèse de lipides bactériens, ce qui persiste dans la synthèse du cholestérol (Microaerobic steroid biosynthesis and the molecular fossil record of Archean life 2011).
4. Un autre intérêt du transfert des gènes mitochondriaux vers le génome nucléaire permet une recombinaison génétique (une fois intégrés au génome nucléaire), ce qui n'est pas possible lors de fission mitochondriale, processus asexué indépendant du cycle cellulaire : ce processus permet l'élimination des mutations défavorables. Mais pourquoi, seulement une partie des gènes mitochondriaux a été transféré vers le noyau dans l'évolution des Eucaryotes ?
- Les différences entre le code génétique de l'hôte et celui de l'endosymbiote pourrait être une des explication, i.e. une la séquence nucléotidique transférée de la mitochondrie vers le noyau ne donnera pas forcément la même protéine, et donc conduirait à la perte de sa de fonction ;
- La grande hydrophobicité de certaines protéines pourrait entraver leur passage à travers les membranes des organites, rendant le transfert de gène difficile (Forces maintaining organellar genomes: is any as strong as genetic code disparity or hydrophobicity? 2005).
- Ces problèmes sont résolus en partie par la fusion mitochondriale.

Par contre, un autre problème survient lors du transfert d'une partie du génome de l'endosymbiote et pose la question de l'origine du noyau.
Origine du noyau
Les introns, i.e. portion d'un gène transcrite en ARN, au sein d'un ARN précurseur, et qui est ensuite éliminée par un processus d'excision programmé (épissage) et qu'on ne retrouve donc pas dans l'ARN mature, sont entrés dans la lignée eucaryote par transfert de gènes de l'endosymbiote mitochondrial à un hôte archéal.
- ils se propagent ensuite vers de nombreux sites des chromosomes hôtes.
- La preuve de ceci est l'observation qu'environ la moitié des introns dans les gènes eucaryotes sont archaïques, présents à des positions conservées tout au long des lignées eucaryotes divergentes, i.e. se trouvant déjà chez l'ancêtre commun des eucaryotes (Introns and the origin of nucleus–cytosol compartmentalization 2006).
Or, l'épissage est lent, de l'ordre de plusieurs minutes par intron (In Vivo Kinetics of mRNA Splicing and Transport in Mammalian Cells 2002), alors que la traduction (translation en anglais), i.e. étape de synthèse des protéines par les ribosomes, à partir de l'information génétique contenue dans les ARN messagers, est rapide, de l'ordre de dix liaisons peptidiques par seconde.
- À ce stade, le cytosol de l'hôte est encore un compartiment procaryote, en ce sens que la traduction co-transcriptionnelle peut y avoir lieu, avec des ribosomes actifs synthétisant des protéines sur les transcrits naissants (5 de la figure ci-contre).
- Cela signifie que les introns sont traduits, conduisant à une expression génique défectueuse à des centaines de locus simultanément, une condition mortelle pour l'hôte, sauf si elle est immédiatement corrigée.
Il existe un nombre fini de solutions à ce problème, outre la machinerie spécifique des eucaryotes, la dégradation des ARNm non-sens (NMD, Nonsense-Mediated Decay).
La NMD est un mécanisme de contrôle de qualité des ARN messagers qui vise à éliminer ceux qui comportent un codon stop prématuré, résultant soit d'une erreur de transcription, soit d'une mutation, soit encore d'une erreur d'épissage (Complex Spliceosomal Organization Ancestral to Extant Eukaryotes 2005).
1. Il suffirait de supprimer tous les introns dans les chromosomes.
Ce processus n'a pas eu lieu, car les positions d'introns sont archaïques (Remarkable Interkingdom Conservation of Intron Positions and Massive, Lineage-Specific Intron Loss and Gain in Eukaryotic Evolution 2003 et Rates of intron loss and gain: Implications for early eukaryotic evolution 2005).
2. Une autre solution serait d'inventer un splicéosome, i.e. particule d'épissage (en anglais, splicing), complexe dynamique de particules ribonucléoprotéiques (composées d'ARN et de plus de 200 protéines) et localisé dans le noyau des cellules, beaucoup plus rapide que les ribosomes.
Le splicéosome moderne a eu plus d'un milliard d'années pour affiner sa fonction, mais il n'est pas devenu plus rapide pour autant.
3. Une dernière solution consisterait à séparer physiquement, par voie spatio-temporelle, le lent processus d'épissage du processus rapide de traduction afin que le premier puisse se terminer avant que celui-ci ne s'installe (6 et 7 de la figure ci-dessus).
(Figure : vetopsy.fr)
La séparation cellulaire implique généralement des membranes, et c'est le principe central de cette hypothèse : la pression initiale qui a conduit à la sélection de la membrane nucléaire était d'exclure les ribosomes actifs de la chromatine active.
- Le processus lent d'épissage peut se terminer autour des chromosomes, permettant ainsi une diffusion plus tardive par l'exportation spécifique des ARNm traités dans le cytosol pour leur traduction.
- Le complexe de pores nucléaires intervient dans la translocation des protéines et de l'ARNm entre le cytosol et le noyau.
La génomique comparative des protéines du complexe des pores nucléaires et des protéines qui composent le nucléole montre que beaucoup d'entre eux partagent des domaines avec des protéines archéennes et bactériennes (Comparative Genomics, Evolution and Origins of the Nuclear Envelope and Nuclear Pore Complex 2004).
Selon ce point de vue, l'origine du noyau marque l'origine d'un compartiment cellulaire véritablement nouveau, non le noyau lui-même, mais le cytosol eucaryote, exempt de chromatine active.
- Les interactions protéine-protéine, plutôt que les interactions protéine-ADN, se déplacent dans le cytosol à des fins de la signalisation et de régulation.
- Les protéines peuvent spontanément s'agréger et interagir de manière à générer de nouvelles structures et fonctions, comme un cytosquelette vrai et les processus de circulation membranaire qui distinguent les eucaryotes des procaryotes.
Ce modèle nécessite seulement que les eucaryotes possèdent une membrane nucléaire lorsqu'ils expriment des gènes, et explique un autre caractéristique sous-estimée qui sépare les eucaryotes des procaryotes.
- Les procaryotes expriment leurs gènes continuellement pendant la division cellulaire, alors que les eucaryotes ont arrêté l'expression de tous leurs gènes avant la division des chromosomes et la division cellulaire.
- Cela suggère un lien évolutif entre l'origine épissage-dépendante du noyau, l'origine des mécanismes génomiques du génome (Epigenetics: Core misconcept 2013) qui impliquent généralement des modifications chimiques de la chromatine et des histones, et l'origine du cycle cellulaire eucaryote.
Cet ensemble d'événements conduit à une cellule bipartite qui (8 de la figure ci-dessus) :
- nécessite un noyau pour exprimer des gènes ;
- a conservé des ribosomes archéaux dans le cytosol en tant que vestiges de l'hôte ;
- possède des bactéries le métabolisme énergétique à la fois dans le cytosol et dans la mitochondrie ;
- a perdu toutes les fonctions de phosphorylation par transfert d'électrons dans la membrane plasmique ;
- a néanmoins conservé l'ATPase archaïque, qui fonctionne maintenant pour acidifier la vacuole ;
- a des caractéristiques eucaryotes typiques.

Les nombreuses théories endosymbiotiques oublient que l'origine des nouveautés des cellules eucaryotes avait un prix énergétique, et ce prix était payé par les mitochondries (The energetics of genome complexity 2010).
- L'internalisation des membranes bioénergétiques chez les eucaryotes les libère des contraintes bioénergétiques qui maintiennent les procaryotes dans leur organisation.
- Cela met la symbiose mitochondriale au tout début de l'eucaryogenèse.
Biologie cellulaire et moléculaireConstituants de la celluleNoyauCytoplasmeCytosolCytosqueletteRéticulum endoplasmiqueAppareil de GolgiMitochondriesLysosomesEndosomesPeroxysomesMatrice extracellulaireReproduction cellulaireBiochimieChimie organiqueBioénergétiqueComposition de la matière vivanteTransport membranaireMoteurs moléculairesVoies de signalisation