Les porines sont des protéines transmembranaires qui laissent passer dans les deux sens des substances de faible poids moléculaire, moins de 5 à 7 kDa.
Hiroshi Nikaido
Leur découverte récente, à la fin des années 1970, est due à Hiroshi Nikaido qui travaillait sur la résistance bactérienne (The porologist 2005).
Remarque : les aquaporines sont des protéines d'une famille plus large de protéines intrinsèques majeures qui forment des pores dans la membrane des cellules biologiques, facilitant principalement le transport de l'eau entre les cellules.
Les aquaporines sont très bien décrites dans l'article de Wikipedia.
Chez les mammifères, les aquaporines sont nombreuses et les plus courantes sont AQP1, AQP2, AQP3 et AQP4.
Vue d'ensemble
Les porines sont retrouvées dans la membrane externe :
Les porines forment des canaux de grande taille qui, contrairement aux autres canaux, laissent passer des molécules non spécifiquement par diffusion facilitée, i.e. transport passif qui ne consomme pas d'énergie.
Porine des glucides
(Figure : vetopsy.fr d'après GFDL, Zephrys et Lomize)
La superfamille I en comprend le plus grand nombre avec 47 familles de porines différentes de bactéries gram-négatives et une famille de porines mitochondriales eucaryotes.
La superfamille IV [Tim17/OEP16/PxMPL (TOP)] contient de nombreux composants des translocases d'organites eucaryotes.
La superfamille III dérive des plastides (comme les chloroplates) des algues, des plantes et de quelques organismes eucaryotes.
Les superfamilles II et V (MspA et Corynebacterial PorA/PorH respectivement) proviennent d'actinobactéries (bactéries gram-positives).
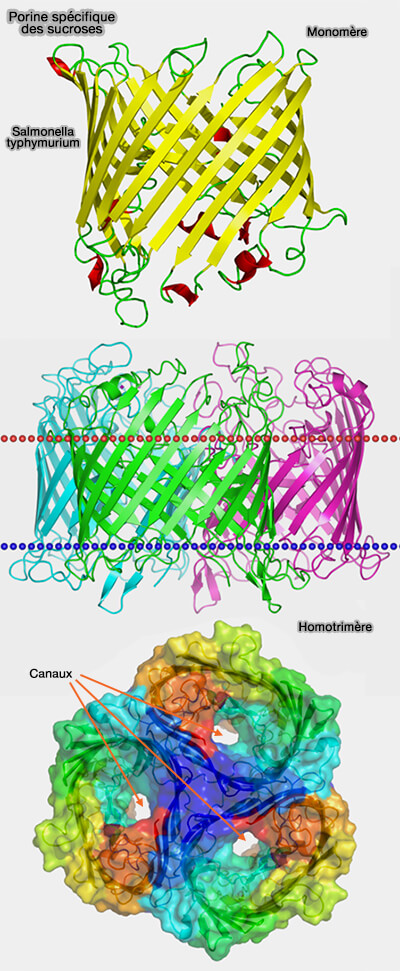
Les porines sont des assemblages de brins β disposés de manière antiparallèle pour former un tonneau β (β barrel) cylindrique, sans fond, qui traverse la membrane externe de part en part (Bacterial and Eukaryotic Porins 2004).
1. Les brins β (β-strand), dont le nombre peut varier de 8 à 22 et qui forment un feuillet β (β-sheet), sont reliés entre eux par :
2. Les brins β sont composés alternativement de résidus polaires et non polaires.
Les résidus non polaires (hydrophobes), orientés vers l'extérieur, se fixent solidement aux lipides membranaires.
Les résidus polaires (hydrophiles), tournés vers l'intérieur du tonneau, créent un milieu aqueux où l'on peut trouver, selon la taille de la porine :
uniquement de l'eau,
deux ou des brins β repliés qui forment une boucle qui détermine la taille des solutés qui peuvent la traverser, comme dans les porines OmpF (Outer Membrane Protein F).
3. Toutes les porines forment des homotrimères, i.e. trois canaux reliés par des liaisons hydrogène et des interactions dipôle-dipôle pour éviter leur dissociation.
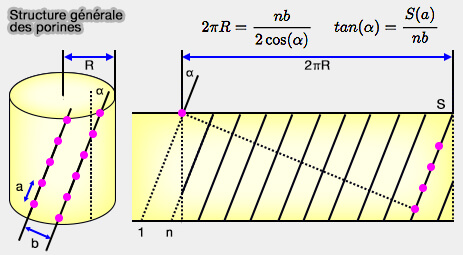
On peut utiliser plusieurs paramètres reliés par des formules mathématiques (Bacterial and Eukaryotic Porins 2004) pour déterminer la structure d'une porine :
$2\pi R=\dfrac{nb}{2\cos(\alpha)}$ et $tan(\alpha)=\dfrac{S(a)}{nb}$ où :
α est l'angle d'inclinaison que forme la porine par rapport à la membrane (entre 30 à 60°),
S, le nombre de " cisaillement " (" shear "), nombre de résidus d'acides aminés trouvés dans chaque brin β,
Structure générale des porines
(Figure : vetopsy.fr d'après Benz)
n, le nombre de brins β de la porine,
R, le rayon du tonneau,
a (3,3 Å) et b (4,4 Å), les paramètres du peptide.






 membrane externe mitochondriale et canaux),
membrane externe mitochondriale et canaux),
{kind=link}
{kind=link}
{kind=link}