Alors que les P1-P3 ATPases sont des transporteurs d'ions, les P4 transportent des lipides volumineux, jusqu'à 10 fois plus gros que les ions des pompes ioniques Ca++-ATPases et Na+/K+-ATPases à structure et topologie similaires.
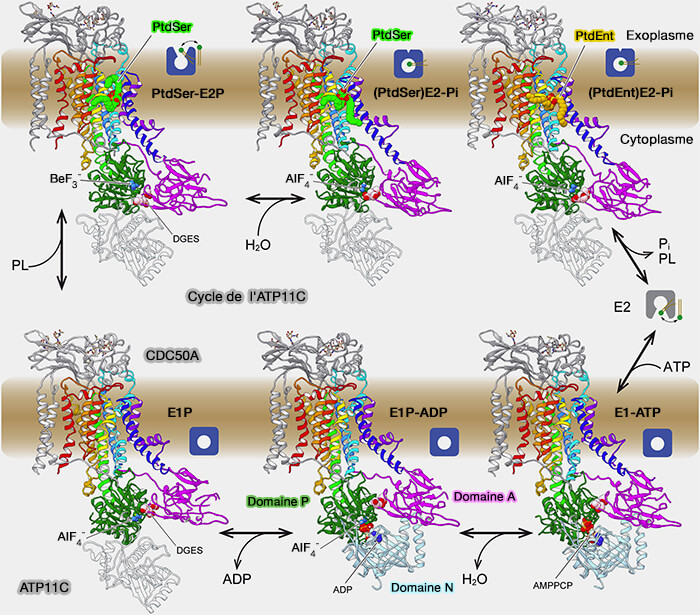
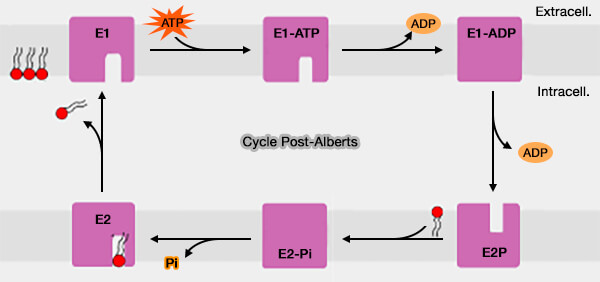
1. Le cycle de transport des P4-ATPases est représenté par le schéma Post-Albers, i.e. à l'état E1 le site de liaison au substrat de transport fait face au côté cytosolique de la membrane tandis qu'à l'état E2, il fait face au côté luminal/extracellulaire.
Cycle Post-Alberts
(Figure : vetopsy.fr d'après Cheng et coll)
2. Le mécanisme, par la structure et la topologie des P4-ATPases plaident en faveur d'un mécanisme de flippase lié à celui connu des pompes ioniques Ca++-ATPases et Na+/K+-ATPases.
Dans le demi-cycle E1 de SERCA ou Na+/K+-ATPase, l'ATP est lié par le domaine N et l'autophosphorylation d'un aspartate invariant au domaine P est étroitement couplée à la liaison et à l'occlusion de deux ions Ca++ ou trois ions Na, respectivement.
La conversion ultérieure à l'état E2P ouvert vers l'extérieur à faible énergie complète l'exportation.
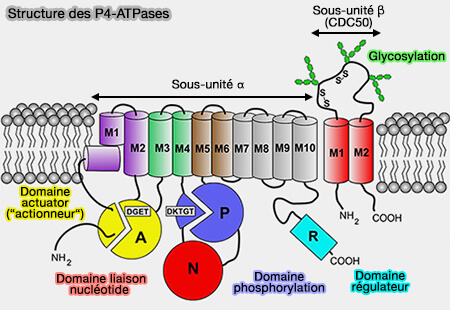
Structure des P4-ATPases
(Figure : vetopsy.fr d'après Andersen et coll)
La liaison d'un substrat de transport extracellulaire ou luminal, i.e. deux ions K+ pour la Na+/K+-ATPase ou un lipide pour les P4-ATPases, est couplée à une auto-déphosphorylation catalysée par une boucle du domaine A avec un motif GE, glycine/glutamate, invariant et un retour ultérieur à l'état E1.
3. Par contre, les P4-ATPases sont fondamentalement différentes des ATPases de type P transportant des ions car leur activité de transport est uniquement associée à la réaction de déphosphorylation, alors qu'aucun transport n'a été démontré pour la phosphorylation
4. Une autre différence clé entre les P4-ATPases et les autres ATPases de type P réside dans la boucle de déphosphorylation du domaine A.
Les ATPases de type P des sous-familles P1-P3 et P5 ont un motif TGES conservé dans le domaine A, où le glutamate est responsable du positionnement d'une molécule d'eau pour attaquer l'aspartate (D) phosphorylé dans le domaine P, et la thréonine stabilise le structure locale de la boucle.
Dans les P4-ATPases, cependant, la séquence est DGET, avec des implications mécanistes encore à expliquer.
Modèle de transport des phospholipides pour l'ATP11C
(Figure : vetopsy.fr d'après Nakanishi et coll)
5. Les structures récentes des P4-ATPases ont fourni de nouvelles informations sur le mécanisme de transport unique des P4-ATPases qu'on peut résumer comme suit ( documentation web).
a. Le lipide substrat est lié avec son groupe de tête dans un sillon dans le domaine TM, dont l'ouverture peut être sujette à l'autorégulation.
Dans l'état E2P ouvert vers l'extérieur, le phospholipide pénètre de la surface du feuillet externe vers la cavité exoplasmique en changeant son orientation.
Le groupe de tête phospholipidique piégé diffuse le long de la crevasse membranaire avec ses queues hydrophobes s'étendant jusqu'au noyau hydrophobe de la bicouche. Le groupe de tête phospholipidique traverse la cavité exoplasmique chargée positivement jusqu'au site d'occlusion près de Val-357 avec TM4.
b. L'occlusion du lipide lié dans le site de transport membranaire est couplée au positionnement du domaine A avec le motif DGET pour la stimulation de la déphosphorylation.
Cycle de phosphorylation de D409 de ATP8A1
(Figure : vetopsy.fr d'après Hiraizumi et coll)
c. La configuration des domaines cytoplasmiques change beaucoup moins pendant la phosphorylation que dans les autres ATPases de type P.
Les liens entre le domaine TM et le domaine A sont plus rigides que dans les autres ATPases de type P et permettent uniquement au domaine A d'effectuer des mouvements de glissement.
Le segment TM4-domaine P est flexible et empêche le mouvement du domaine P lors de la phosphorylation pour provoquer des changements conformationnels dans le domaine TM.
Le cycle pourrait se déroulera comme suit.
1. La liaison de l'ATP à E1 décale légèrement le domaine N, mais ne provoque pas autrement de changements conformationnels à grande échelle, lors du passage à l'état E1-ATP.
2. La phosphorylation d'Asp560 dans le domaine P déplace le domaine N vers le haut, donnant l'état E1P-ADP. Cela déplace à son tour légèrement le domaine A vers l'extrémité N-terminale de Cdc50p, qui interagit également avec les segments M4-P, le stabilisant probablement légèrement.
Modèle du cycle de transport des P4-ATPases
(Figure : vetopsy.fr d'après Timcenko et coll)
3. La libération d'ADP permet au domaine A de se déplacer vers le site de phosphorylation, entraînant avec lui TM1-2 en raison des lieurs rigides du domaine A. Cela ouvre la voie de transport des lipides vers la lumière, exposant TM4 et entraînant l'état E2P.
4. La liaison du substrat, ici PS, entraîne la fermeture du site d'entrée, déplaçant davantage le domaine A lors de la déphosphorylation.
5. La libération de PS et Pi ramène le complexe à l'état E1.
Drs2p interagit avec PI(4)P à travers ce domaine et cette interaction est nécessaire pour l'activité de la flippase de la phosphatidylsérine (PS).
Le domaine de liaison PI4P dans le domaine C-terminal de Drs2p chevauche le site de liaison d'un autre élément régulateur de l'activité flippase, la protéine Arf-GEF Gea2p.
a. L'activité de Drs2p est abolie en l'absence d'interactions avec Gea2p ou de PI(4)P.
Complexe trimère Drs2p, Gea2p et Arl1p
(Figure : vetopsy.fr d'après Tsai et coll)
Le domaine C-terminal de Drs2p agit comme un domaine autoinhibiteur en interférant avec les domaines catalytiques de la protéine.
L'hélice amphipathique se propage dans une direction contraire à la partie restante du domaine auto-inhibiteur et exerce une force de traction mécanique qui détend les interactions entre le domaine auto-inhibiteur et le segment hélicoïdal de queue H1C avec le domaine P ( mécanisme).
Gea2p, Arl1p et Drs2p forment un complexe protéique qui affecte la dynamique membranaire et l'asymétrie lipidique du Golgi.
a. La liaison d'Arl1p aux régions N-terminales et de Gea2p aux régions C-terminales de Drs2p stimule l'activité flippase dépendante de l'ATP de Drs2p au niveau de la membrane du réseau trans-Golgi (TGN).
Le Drs2p actif augmente la surface du feuillet cytosolique aux dépens du feuillet luminal et plie la membrane vers le cytosol.
Ensuite, la golgine Imh1p est capable d'interagir avec la membrane et conserve sa localisation au Golgi.
La suppression de GEA2 ou DRS2 altère la formation du complexe protéique Arl1-Drs2-Gea2, ce qui affecte l'asymétrie PS du TGN et la localisation au Golgi de Imh1p.
b. La formation de ce complexe trimérique est nécessaire à l'activité flippase et au recrutement de la cargaison lors de la formation des vésicules au TGN.
3. Alors que Drs2p interagit avec Gea2p au niveau du TGN, il interagit avec la protéine F-box Rcy1p au niveau d'un domaine chevauchant dans les endosomes précoces.
Les deux interactions différentes de Drs2p avec Rcy1 et Gcs1 au niveau des endosomes précoces peuvent être nécessaires au recrutement de différents types de molécules adaptatrices lors de la formation de vésicules de recyclage destinées à différentes membranes.
Mécanisme d'autorégulation de Drs2p-Cdc5Op
Le mécanisme d'autorégulation de Drs2p-Cdc50p peut être modélisé.
E2Pinhib a un domaine TM fermé avec les domaines cytosoliques verrouillés par le C-terminal auto-inhibiteur, i.e. l'enroulement étendu de l'extrémité C-terminale autour des domaines cytosoliques A, P et N. Phe-1275 et Phe-1277 occupent la poche de liaison à l'ATP, empêchant l'accès de l'ATP au site catalytique.
1. Le déplacement du phospholipide allostérique (PL) par PI4P déclenche la formation de l'hélice amphipathique du côté C-terminal de TM10, qui a deux effets.
Elle provoque le dépliage partiel du reste de l'extrémité C-terminale, déstabilisant ainsi son interaction avec le domaine P par le déplacement de la queue H1C avec le site de liaison supposé à Gea2p.
Elle forme un site d'interaction pour la formation de la boucle TM6-7, qui se déplace ensuite avec le domaine P.
Ensemble, ces deux mouvements déplacent les domaines P et N vers l'hélice amphipathique en grande partie comme un corps rigide.
Autorégulation de Drs2p/Cdc50p
(Figure : vetopsy.fr d'après Timcenko et coll)
2. Le déplacement de l'extrémité C-terminale des domaines cytosoliques entraîne un léger mouvement des domaines N et A, qui amorce l'ouverture du domaine TM par un mouvement de TM1-2 pour la liaison lipidique du substrat, ce qui donnerait lE2Pinter.
3. Le déplacement complet de l'extrémité C-terminale conduit à l'état E2Pactif où une nouvelle rotation des domaines N et A entraîne l'ouverture d'un site supposé de liaison de substrat par le mouvement de TM2 loin de TM6.
Remarque : une protéine auxiliaire, par exemple, Gea2p, pourrait séquestrer l'extrémité C-terminale pour empêcher le lipide de se lier de nouveau.
Autres régulations de Drs2p-Cdc5Op
L'activité des P4-ATPases est également régulée par la phosphorylation et la concentration des sphingolipides.
Sans elles, l'absorption par la membrane plasmique des phospholipides est abolie bien que les flippases soient localisées normalement sur la membrane.
L'activité de Fpk1 et Fpk2 est en outre régulée par la kinase, Ypk1p.
Par conséquent, la phosphorylation catalysée par Ypk1 de Fpk1/2 élimine l'activité de ces protéines, et l'activité de Ypk1 est à son tour inhibée en raison de la phosphorylation par les protéines Fpk1/2.






 documentation web).
documentation web).