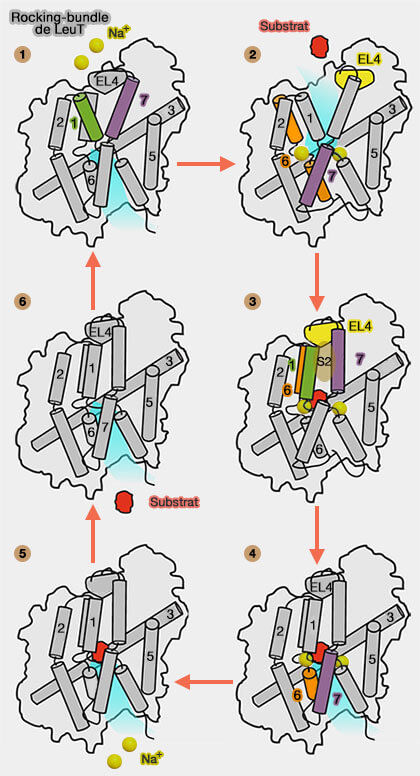
Le mécanisme exact du modèle rocking-bundle (faisceau à bascule) des co-transporteurs est encore incertain et plusieurs modèles de déclenchement ont été proposés.
Il est encore difficile de déterminer si les observations montrent bien des portes réelles ou des artefacts cristallins.
Plusieurs états conformationnels ont été capturés dans les structures cristallines de LeuT et de dDAT.
Mécanisme simplifié de rocking-bundle de LeuT
(Figure : vetopsy.fr d'après Kazmier, et coll)
Toutefois, chaque structure ne représente qu'un instantané dans le cycle de transport, et la transition d'un état à un autre ne peut pas être déduite d'une manière précise.
LeuT
Mécanisme du rocking-bundle
LeuT, sans liaison au substrat et aux ions, alterne entre les conformations ouvertes vers l'extérieur et vers l'intérieur.
2. TM1b, TM6a et TM7 du domaine central se déplacent pour assurer l'occlusion du site de liaison au substrat, qui se traduit par une conformation occluse vers l'extérieur.
Le rapprochement entre les deux domaines, via EL4 (entre les segments TM7 et TM8), ferme la porte extérieure.
Ces événements structuraux semblent précéder le mouvement de bascule du faisceau central qui conduit à une conformation occluse vers l'intérieur.
3. Une fermeture complémentaire associée à EL4 et TM5 a lieu pour provoquer le changement de conformation vers l'intérieur.
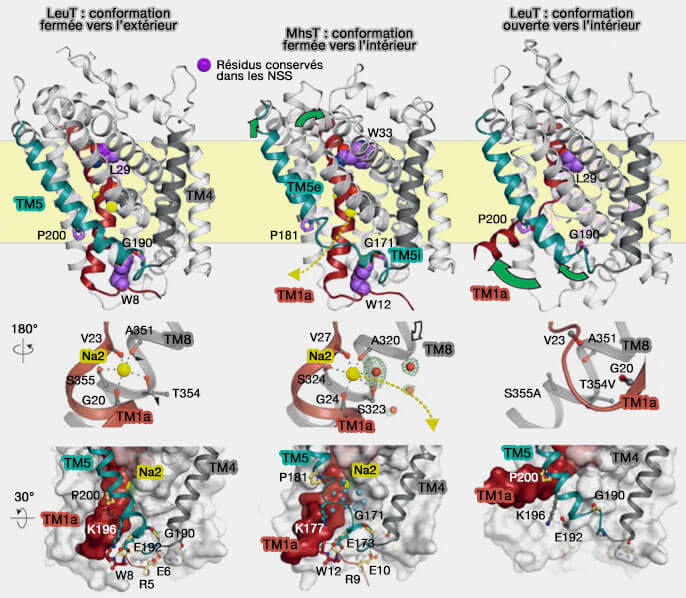
Dans cette conformation, TM5e s'est déplacé par rapport à TM5i, qui, lui, maintient son interaction avec le domaine N-terminal et avec TM1a par la coordination conjointe de Na+ sur le site Na2.
TM5 s'étend donc comme un " ressort " à l'interface de la membrane cytoplasmique, le déroulement étant facilité par les points de déformation GlyX9Pro conservés.
Le déroulement du TM5 fournit un accès du solvant intracellulaire au site Na2, le site Na2 acceptant une molécule d'eau en tant que sixième ligand.
4. La translocation du substrat est couplée à la libération du sodium de Na2.
Na2 libéré, éventuellement stimulé par la flexibilité structurelle de TM5 et un potentiel membranaire négatif, TM1a se libère de TM6 et TM5i se réassocie avec TM5e dans la reformation de l'hélice TM5.
Ces modifications provoquent l'ouverture de la porte et la libération du substrat et de Na1 dans l'environnement intracellulaire.
Mécanisme de " ressort " de TM5
(Figure : vetopsy.fr avec l'aimable autorisation de M. Quick)
1. Des variations du système décrit ci-dessus peuvent être observées dans d'autres transporteurs à repliement LeuT qui se déplacent plus ou moins ou dont le déclenchement varie quelque peu, comme :
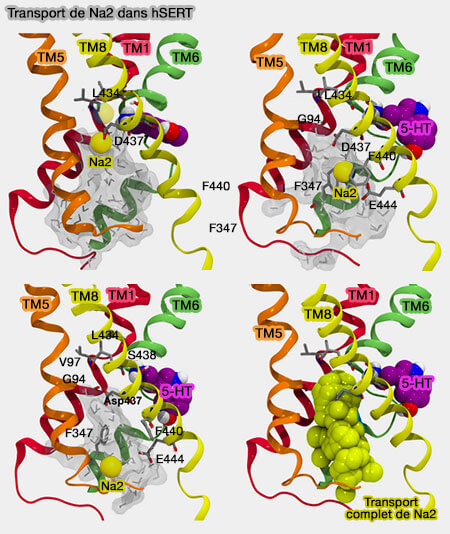
1. Dans les trois MAT, le site Na2 contient un résidu d'aspartate essentiel dans l'ouverture de la porte intracellulaire.
Pour hSERT, la rotation de la chaîne latérale de D437 libère Na+ du site Na2 et le pousse dans la voie de libération intracellulaire située entre TM1, TM5, TM6 et TM8.
L'ion Na+ dans le site Na2 est à l'origine du transport du substrat, et la libération du sodium se produit avant la libération du substrat.
2. Le changement conformationnel entre les configurations ouvertes vers l'extérieur et celles vers l'intérieur peut être décrit comme un mouvement des hélices du faisceau par rapport à l'échafaudage, comme décrit pour LeuT.
une voie le long de TM6b et de TM8 plus fréquente,
une voie alternative le long de TM1a et TM6b, identique à celle de la libération du Na+, décrite précédemment.
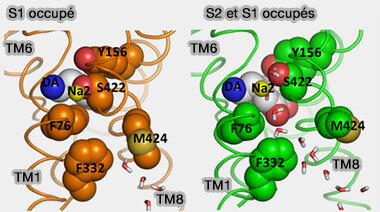
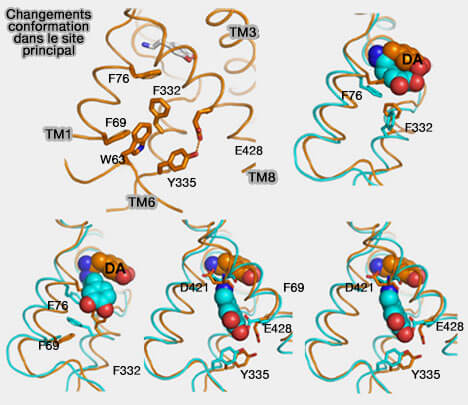
Site vestibulaire de DAT
(Figure : vetopsy.fr d'après Shan et coll)
b. Il semble que des changements rotamériques dans F76 et F322 permettent au DA de se déplacer du site de liaison primaire vers le milieu intracellulaire.
La rotation de F332 provoque le mouvement de F69 qui éloigne TM1a de TM6, suivi de la rupture d'une liaison hydrogène entre Y335 et E428 permettant le mouvement continu de TM1a et l'extrémité N-terminale de TM6.
L'eau serait capable de se déplacer jusqu'au site de liaison primaire quand une molécule DA est localisée dans chacun des deux sites. lorsque la dopamine occupe seulement le site primaire, l'eau accède uniquement à F332, qui est approximativement à mi-chemin du site de liaison primaire.
Site S2 nécessaire ou pas ?
Est-ce que le substrat se lie uniquement au site de liaison primaire de LeuT et MATs ou la liaison d'un second substrat dans le vestibule extracellulaire (site vestibulaire) est nécessaire pour déclencher la libération du substrat du site de liaison primaire ?
Site vestibulaire de DAT
(Figure : vetopsy.fr d'après Shan et coll)
La fonction du site vestibulaire dans les MAT et d'autres transporteurs à repliement LeuT fait donc encore l'objet de débats.
Toutefois, il semblerait que la liaison du substrat dans le site vestibulaire puisse accélérer l'ouverture vers l'intérieur de hDAT.
La molécule DA occupant le site de liaison primaire augmente son interaction avec l'eau lorsque le site vestibulaire est également occupé en raison d'une rotation de S422 et M424 qui permet à l'eau de se déplacer vers le site Na2 du côté intracellulaire.
Avant que DA soit libérée dans le cytoplasme, une interaction de DA complètement solvatée avec E428 semble être la dernière interaction stabilisante entre DA et son récepteur avant sa libération.
Analogies et différences entre les mécanismes rocker-switch et rocker bundle
2. Le site de liaison au substrat situé à l'interface se trouve à mi-chemin de la membrane.
3. Les transitions entre les conformations orientées vers l'extérieur (outward-facing ou outward-open) et celles vers l'intérieur (inward-facing ou inward-open) sont la conséquence des mouvements des domaines transmembranaires les uns par rapport aux autres.
4. La liaison du substrat est associée à des changements conformationnels locaux, impliquant des parties flexibles (non enroulées) des domaines hélicaux.
Par contre, on peut noter certaines différences.
1. Les domaines sont, en général, symétriques dans les rocking-switch, distincts dans les rocking-bundle.
2. Les événements de déclenchement sont bien plus complexes dans les rocking-bundle que dans les rocker-switch (comparaison entre LeuT et GLUT).
2. Les rocker-switch sont plus simples et plus plastiques que les rocking-bundle qui sont eux plus spécifiques et plus diversifiés.
Les transporteurs passifs facilitateurs n'utilisent que des mécanismes simples à bascule.





 site vestibulaire) est nécessaire pour déclencher la libération du substrat du site de liaison primaire ?
site vestibulaire) est nécessaire pour déclencher la libération du substrat du site de liaison primaire ?