a. La classification SCOP est basée sur une orientation différente des huit brins β de chaque groupe de domaines, ce qui n'est d'ailleurs pas un facteur déterminant pour la fonction du domaine, y compris pour leurs propriétés à lier Ca++.
b. Cette superfamille comprend deux familles ( tableau) :
les variantes de type synaptotagmine également appelées famille S ou topologie de type I,
les variantes de type phospholipase C (PLC), également connues sous le nom de famille P ou de topologie de type II,
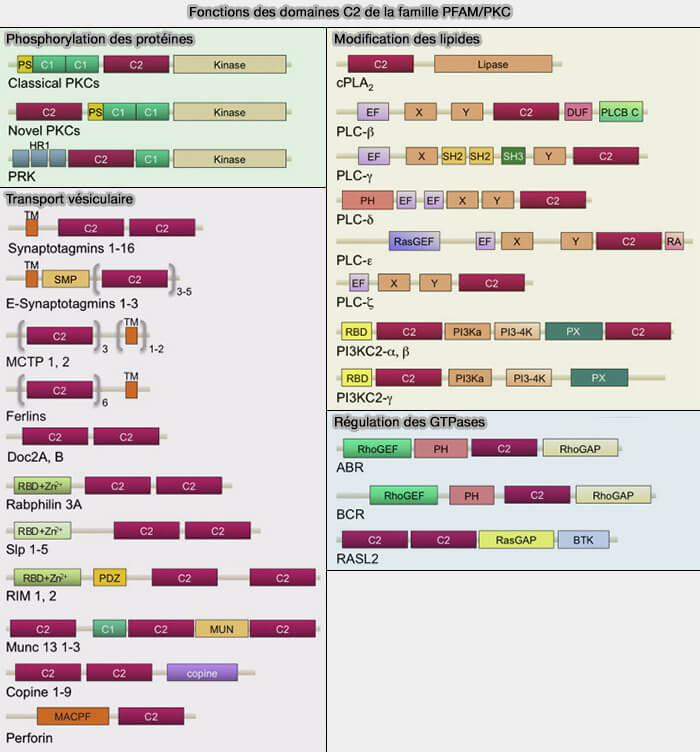
c. La base de données PFAM (Protein Families) classe les domaines C2 en plusieurs groupes ( tableau).
2. Le domaine C2 est retrouvé dans les PKC classiques (α, βI, βII, γ), comme deuxième des quatre domaines conservés responsables de la liaison membranaire dépendante de Ca++.
Les nouvelles PKC (ε, η, δ, θ) n’ont pas cette dépendance au Ca++, mais elles contiennent une région V0/C2 à leur N-terminal qui interagit avec des phospholipides chargés négativement.
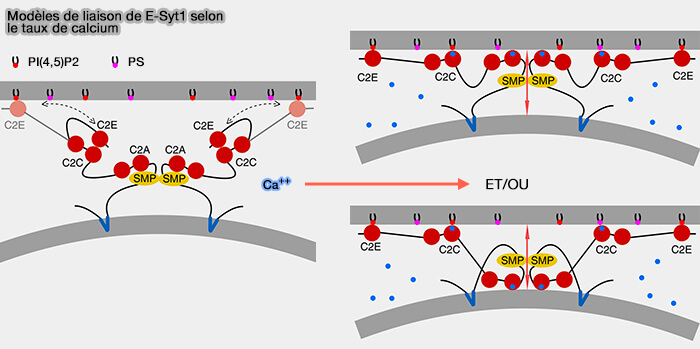
en plusieurs exemplaires dans la même protéine par exemple, E-Syt1-3, protéines de type synaptotagmine ou extended synaptotagmines, régulent aussi la formation des MCS du RE/MP et le transfert des lipides,
Modèles de liaison de E-Syt1 selon le taux de calcium
(Figure : vetopsy.fr
d'après Bian et coll)
avec un ou plusieurs domaines C1,comme dans les PKC.
3. Bien que tous les domaines C2 partagent une structure commune, de légères variations dans les résidus situés dans les boucles d’interconnexion confèrent aux domaines C2 des capacités fonctionnelles :
de répondre à différentes concentrations de Ca++ et de lipides,
de servir de signalisation par des interactions protéine-protéine.
Grâce à ces propriétés, le domaine C2 est un acteur crucial dans de nombreuses fonctions cellulaires.




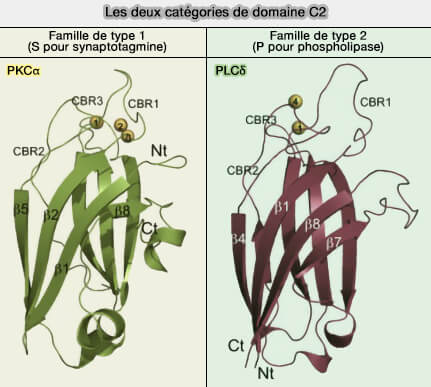
 tableau) :
tableau) :