Les protéoglycanes (PG) sont des glycoprotéines O-glycosylées caractérisées par la présence d'une protéine centrale avec une ou plusieurs chaînes de glycosaminoglycanes (GAG) attachées de manière covalente.
Les PG sont localisés :
à la surface des cellules,
dans la matrice extracellulaire
dans la circulation.
Vue d'ensemble des protéoglycanes
La liaison des glycosaminoglycanes (GAG), i.e. chaînes glucidiques, à un cœur protéique forme un complexe appelé protéoglycane (PG).
1. Les GAG étant classés en quatre groupes, en fonction de leurs structures disaccharidiques centrales, les protéoglycanes peuvent être à base :
Remarque : seul l'hyaluronane ou acide hyaluronique (HA) est retrouvé à l'état libre, sans aucune liaison covalente avec une protéine. En outre, il n'est jamais sulfaté.
2. On trouve plus de quarante coeurs protéiques différents qui ne forment pas seulement des échafaudages.
Leurs domaines peuvent être engagés dans un certain nombre d'interactions différentes et de nombreux noyaux protéiques présentent des variantes résultant d'un épissage alternatif.
Structure des protéoglycanes
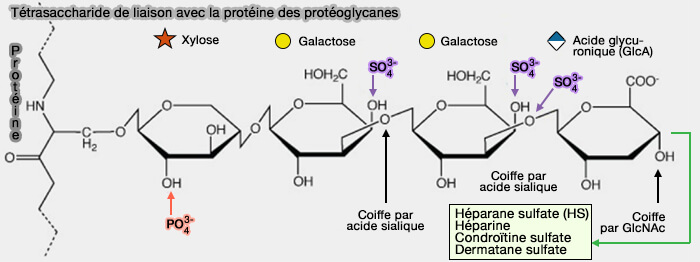
1. Les GAG sont attachés sur le coeur protéique par un résidu sérine (Ser) selon une liaison tétrasaccharidique différente selon les PG.
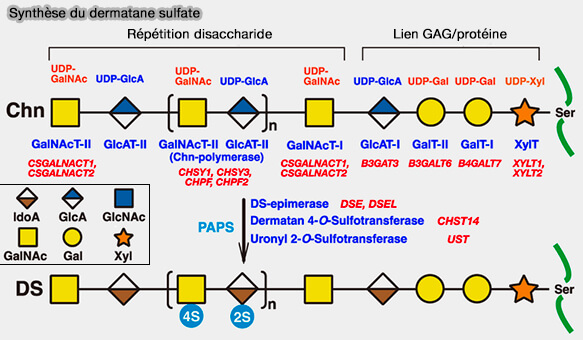
Par exemple, CSPG a pour liaison : acide glucuronique (GlcA) -galactose (GAL)-galactose (GAL)-xylose(Xy)l-Ser, DSPG, IdoA-Gal-Gal-Xyl-Ser, i.e. IdoA remplaçant GlcA par épimérisation sur C5.
Synthèse du dermatane sulfate
(Figure : vetopsy.fr d'après Mizumoto et coll)
Le résidu Ser est généralement dans la séquence -Ser-Gly-x-Gly- (où x peut être n'importe quel résidu d'acide aminé sauf la proline).
2. Les chaînes sont de longs polymères glucidiques linéaires chargés négativement dans des conditions physiologiques en raison de la présence de groupes sulfate et d'acides uroniques.
La sulfatation est essentielle pour de nombreux processus comme :
pour les KSCG, i.e. dans la transmission d'informations importantes sur la reconnaissance moléculaire et le comportement cellulaire à travers un certain nombre de protéines interactives ( kératane sulfate).
3. Les coeurs des protéines PG peuvent influencer le type et les schémas de modification des chaînes GAG attachées par la suite.
Toutefois, la manière dont les informations sont transmises du noyau protéique aux enzymes engagées dans la polymérisation et la modification des GAG est obscure.
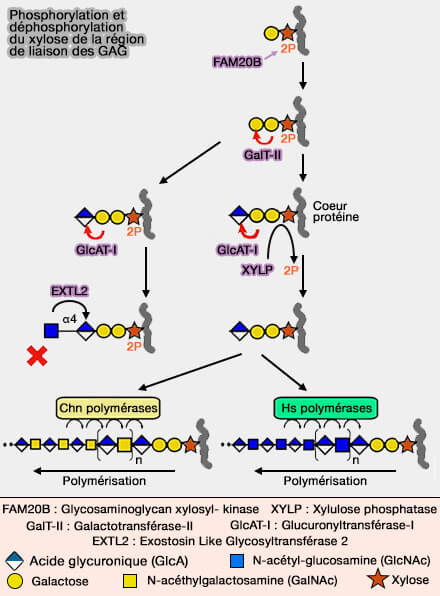
Un mécanisme suggéré est la sulfatation et la phosphorylation des unités du tétrasaccharide de liaison qui attache les chaînes GAG à un site de modification d'un noyau protéique.
Par contre, comme un même noyau protéique peut acquérir différents types de chaînes GAG dans différents types de cellules, la modification des GAG pour un noyau protéique particulier pourrait être spécifique à un type de cellule et de tissu.
Par contre, on ne sait rien du comment et du pourquoi de ces modifications.
3. Le tétrasaccharide de liaison peut subir des modifications dont on ne sait pas encore si elles ont lieu dans les sites de sortie du réticulum endoplasmique (RE) ou dans l'appareil de Golgi.
4. La sulfatation de la région de liaison est limitée aux voies CS et DS, i.e. le deuxième galactose est 4-O-sulfaté et les première et deuxième unités de galactose sont 6-O-sulfatées.
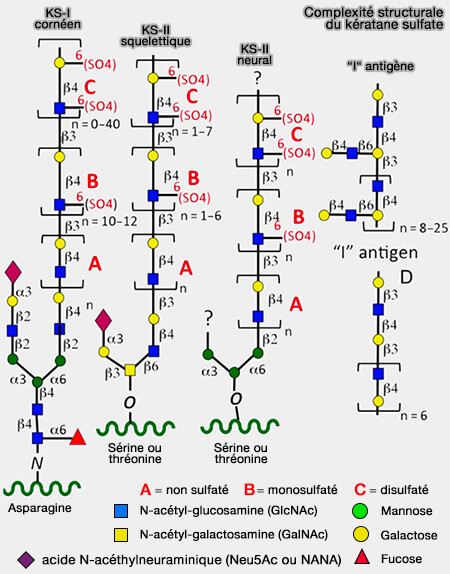
Les KSPG sont les seuls PG dans lesquels les chaînes de kératane sulfate (KS) ne sont pas attachées au noyau protéique via le tétrasaccharide de liaison.
Complexité structurale du kératane sulfate
(Figure : vetopsy.fr d'après Caterson et coll)
1. Les différents modes d'attachement des chaînes KS à leurs noyaux protéiques dans les KSPG constituent la base de la distinction entre KSI, KSII et KSIII.
a. Pour KSI, les chaînes GAG sont liées au noyau protéique via une structure de glycane complexe liée à l'asparagine (N-glycosylation).
Une antenne en C6, et parfois deux en C3 et C6 peuvent être aussi modifiées.
c. Les chaînes KS de type III sont abondantes dans le cerveau et sont liées au noyau protéique via la fixation du mannose aux résidus de sérine ou de thréonine.
dans les HA
L'acide hyaluronique (HA), GAG non sulfaté sans liaison protéique covalente, est abondant dans les matrices extracellulaires et à la surface des cellules.
Toutefois, HA est souvent lié à des PG appelés lecticanes qui possèdent un domaine de liaison HA appelé protéine de liaison.






 kératane sulfate).
kératane sulfate).