La technique du potentiel stabilisé ou potentiel imposé (voltage clamp) permet d'interpréter les phénomènes qui ont lieu lors du déroulement d'un potentiel d'action.
La membrane se comporte comme un circuit avec un condensateur et une résistance placée en parallèle.
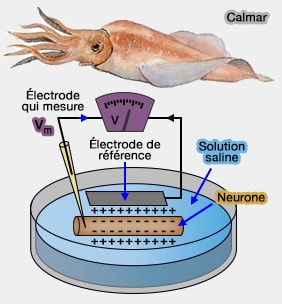
Dans cette technique qui est la plus simple, on plonge une cellule, au départ un axone géant de calmar, dans une solution saline et un voltmètre est relié à la préparation au moyen d'électrodes.
Mesure du potentiel de repos
(Figure : vetopsy.fr)
Le voltmètre est en général composé de tubes capillaires en verre dont la pointe de moins de 1 micron peut être introduite dans la cellule sans la léser gravement.
Les électrodes contiennent une solution saline conductrice KCl3 molaire : la vitesse de diffusion des ions est identique et ne fausse pas la mesure.
L'axone de Calmar a été choisi car :
son diamètre peut atteindre 1 mm, soit 100 à 1000 fois celui des axones des mammifères, d'où le nom " géant " ;
1. Les deux électrodes se trouvent dans le milieu extracellulaire : aucune différence de potentiel n'est enregistré. Le potentiel du milieu extracellulaire est en général pris comme " zéro " pour les mesures ultérieures.
2. On introduit la pointe de l'électrode dans la cellule en perforant sa membrane : la différence de potentiel est de 70mv.
Comme l'intérieur de la cellule est négatif par rapport à l'extérieur, la valeur du potentiel de repos est de - 70 mV.
Ce potentiel dit de repos dépend du type de neurone et varie entre -40 à -90 mV.
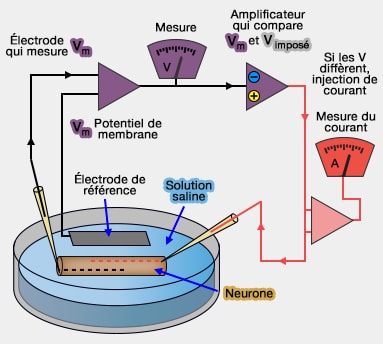
Voltage clamp
(Figure : vetopsy.fr)
Voltage imposé ou voltage clamp
Cette technique du potentiel stabilisé ou potentiel imposé (voltage clamp) permet de soumettre la membrane à des échelons de dépolarisation ou d'hyperpolarisation qui permettent d'interpréter les phénomènes qui ont lieu lors du déroulement d'un potentiel d'action.
Le stimulus artificiel le plus efficace pour déclencher un potentiel d'action est un choc électrique, et suivre l'évolution de la perméabilité de la membrane aux ions revient à mesurer les modifications de la résistance membranaire (ions incriminés) par la loi d'Ohm ($V=RI$).
Si on parvient à faire passer dans la membrane un courant tel que la dépolarisation membranaire passe de façon abrupte de son niveau de repos à un niveau imposé et stabilisé (dépolarisation en échelon), le potentiel de la membrane reste alors stable.
$dV/dt=0$, donc $I_c$, ce qui implique que $I_t=I_r$.
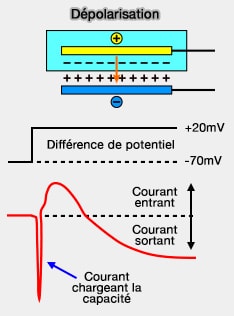
courant entrant, i.e. les charges positives (cations) entrent dans la cellule,
courant sortant, i.e. les charges positives sortent de la cellule.
Remarque importante : par convention, le courant entrant est représenté par une courbe orientée vers le bas, un courant sortant par une courbe vers le haut. Dans la figure ci-contre, le sens est inversé.
Appareillage
L'élément essentiel de l'appareillage du voltage stabilisé consiste en un générateur de courant contrôlé par une rétroaction.
Le courant est débité par l'intermédiaire de deux électrodes : l'une est insérée dans l'axoplasme, l'autre est extracellulaire. Le courant injecté est mesuré de façon permanente par un appareil adéquat.
Une deuxième paire d'électrodes (l'une intra, l'autre extracellulaire) est branchée à un appareil de mesure de la différence de potentiel.
On fixe sur le générateur de courant le niveau de polarisation auquel on désire que la membrane soit soumise (potentiel stabilisé).
Le générateur de courant à rétroaction compare sans arrêt le niveau de la différence de potentiel désirée à la valeur de la différence de potentiel mesurée à travers la membrane.
Et si les deux valeurs ne sont pas égales, il modifie l'intensité du courant qu'il débite pour qu'elles le deviennent.
Décomposition K+ et Na+
(Figure : vetopsy.fr)
Contributions respectives des ions
1. Si on impose à la membrane une dépolarisation de 90mV, on observe la courbe de courant ci-dessus qui s'explique avec l'équation de Nerst.
On observe, tout au départ, le courant qui charge la capacité, qu'on ne représentera plus par la suite.
Puis, la dépolarisation classique de la membrane.
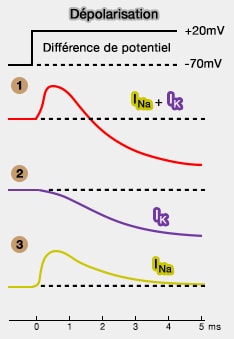
2. On impose à la membrane une dépolarisation de 90mV, mais l'axone baigne dans une solution sans sodium : on trouve la courbe 2 ci-contre.
Le courant entrant qui a disparu correspond au sodium.
Si on soustrait ce courant $I_K$ au courant de départ, on trouve le courant $I_{Na}$.
Perméabilité Na+ et K+
(Figure : vetopsy.fr)
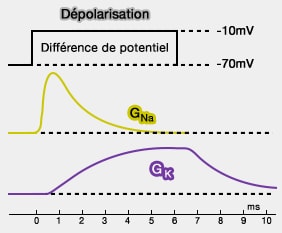
Selon la loi d'Ohm, on peut donc écrire : $G_{Na}=\dfrac{I_{Na}}{V_{imposé}-E_{Na}}$
Or, on connait $V_{imposé}$ car il a été choisi par l'expérimentateur, $E_{Na}$ qui est constant, $I_{Na}$ qui est représenté par la courbe, ce qui permet de définir $G_{Na}$, i.e. la perméabilité de la membrane aux ions Na+.
Les déterminations
expérimentales ont montré que,
pour les cellules nerveuses et les cellules musculaires, $G_K$ est 10 à 25 fois plus grand que $G_{Na}$.
Sur la figure, on voit que :
la perméabilité de Na+ augmente rapidement, puis diminue et revient à son état initial malgré le maintien de la dépolarisation imposée (-10 mV).
La perméabilité de K+ s’accroît plus lentement, mais reste à un palier pendant toute la dépolarisation. Puis, elle diminue progressivement pour revenir lentement à la normale.



{kind=link}


 ions incriminés) par la loi d'Ohm ($V=RI$).
ions incriminés) par la loi d'Ohm ($V=RI$).