


Bases moléculaires de la mémoire à long terme déclarative
Potentialisation à long terme ou PLT (2)
- Généralités sur la mémoire
- Bref historique de la mémoire
- Modèles et définitions de la mémoire
- Différentes mémoires
- Bases
neurobiologiques de la mémoire
- Bases neurobiologiques générales de la mémoire
- Bases neurobiologiques de la mémoire à court terme
- Bases neurobiologiques de la mémoire de travail
- Bases neurobiologiques de la mémoire à long
terme
- Bases anatomiques de la mémoire à long terme non-déclarative
- Bases anatomiques de la mémoire
à long terme déclarative
- Bases anatomiques de la mémoire à long terme déclarative : découvertes
- Bases anatomiques de la mémoire à long terme déclarative : lobes temporaux
- Bases anatomiques de la mémoire à long terme déclarative : diencéphale
- Bases anatomiques de la mémoire à long terme déclarative : lobes frontaux
- Bases anatomiques de la mémoire émotionnelle : système limbique
- Bases moléculaires de la mémoire à long terme
La potentialisation à long terme ou PLT, dans un premier temps, augmente la force et la durée du potentiel d'action dans le neurone postsynaptique.
PLT : synthèse de récepteurs et de dendrites
Mise en évidence
Une expérience célèbre a montré que le passage en mémoire à long terme implique la synthèse de nouvelles protéines.
On apprend à des souris à tourner à droite ou à gauche dans un labyrinthe en T. Certaines souris reçoivent un inhibiteur de la synthèse protéique une heure avant ou tout de suite après l'expérience.
1. Les deux groupes apprennent correctement la tâche et répondent parfaitement peu après, i.e. la mémoire à court terme est bonne.
2. Quand elles sont testées plusieurs heures après :
- les souris témoins répondent normalement ;
- les souris ayant reçu l'inhibiteur ont un déficit très important de cette mémoire à long terme.

La synthèse protéique est donc nécessaire aux changements
durables de la connexion synaptique : la facilitation à long terme
de l'aplysie en est un exemple caractéristique ( sensibilisation à long terme).
sensibilisation à long terme).
- Cette synthèse doit être effectuée pendant l'activité synaptique précise, i.e. dans ce cas, l'application de la sérotonine sur les neurones modulateurs.
- Ce qui explique que les inhibiteurs de la synthèse protéique sont efficaces avant ou juste après l'apprentissage.
Mécanisme d'action
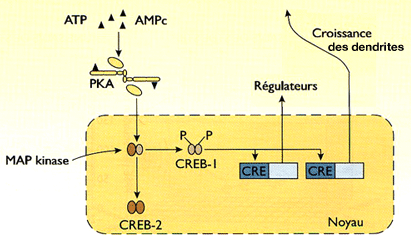
Qui dit synthèse protéique dit activation de gènes !
1. La
CaM kinase II, non seulement s'autophosphoryle et phosphoryle les récepteurs
NMDA (rôle des ions Ca++ ), mais active des MAP kinases, (Mitogen-activated protein kinases ou MAPK).
Squire-Kandel : La mémoire)
Les MAP kinases, par leur action sur les gènes et donc sur les protéines, augmentent la densité des synapses et peuvent, de ce fait, inclure des récepteurs supplémentaires dans leurs membranes.
2. En outre, les unités catalytiques de la PKA permet l'ouverture des canaux
AMPA (PKA et calmoduline).
Si ces stimulations sont nombreuses, ces unités convergent vers le noya.
- Elles phosphorylent un facteur de transcription, la CREB-1 (C-AMP Response Element-binding protein), i.e. protéine se fixant au CRE.
- Le CREB se fixe sur le CRE (cAMP response élément) de l'ADN, qui contient la séquence nucléotidique hautement conservée, 5'-TGACGTCA-3', pour activer les gènes permettant la fabrication de nouveaux récepteurs AMPA (La biologie moléculaire de la mémoire: un dialogue entre gènes et synapses 2000).
3. De nouveaux récepteurs sont incorporés à la membrane dendritique dont la surface a été déjà multipliée par l'action des MAP kinases.

Les protéines fabriquées dans la plasticité synaptique se dirigent uniquement vers les synapses concernées, et non vers toutes les synapses du neurone (environ 1000 par neurone), ce qui explique la spécificité de la mémorisation, aussi bien dans la fabrication que dans la récupération des souvenirs.
(Figure : vetopsy.fr d'après
Sciences magazine)
4. Toutefois, si la CREB-1 est un activateur, la CREB-2 est un répresseur qui se fixe sur le CRE et sur le CREB-1 pour empêcher cette activation.
La régulation de ce dernier est indépendante de la PKA, mais est sous le contrôle d'une MAP kinase.
Ceci pourrait expliquer les différences de passage de la mémoire à court terme à celle à long terme en fonction de ces deux régulations (vieillissement, mémoire exceptionnelle comme Shereshevsky…).
- On observe un doublement (1300 à 2600) des terminaisons synaptiques des neurones présynaptiques chez l'aplysie, mais également une augmentation dendritique des neurones postsynaptiques.
- Lors du déclin de la mémoire, les terminaisons
synaptiques régressent. Dans le cas de l'habituation à long
terme (infos), l'apprentissage aboutit à une diminution du nombre de
synapses (1300 à 800).
PLT et DLT dans les système nerveux central
La PLT et la DLT ont été également découvertes dans le néocortex qui est également un lieu de stockage de la mémoire déclarative à long terme.
Les mécanismes de la plasticité synaptique, lors de l'apprentissage et la mémorisation, ressemblent à ceux du développement embryonnaire cérébral.
Ces phénomènes expliquent
que les souvenirs anciens, comme chez H.M. (découverte de la mémoire à long terme), sont conservés et non nullement besoin de l'aide du lobe
temporal interne.