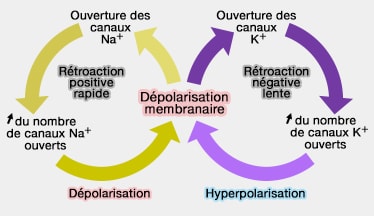
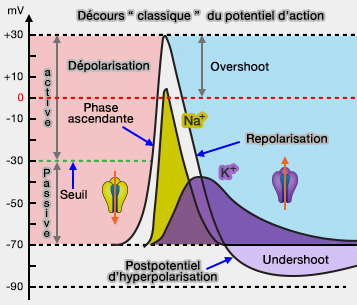
Cette entrée d'ions Na+ provoque une dépolarisation qui ouvre de plus en plus de canaux sodium, ce qui emballe la réaction (dépolarisation active : cycle de Hodgkin).
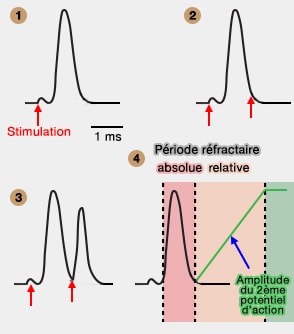
Cette dépolarisation correspond à la phase ascendante d'un potentiel d'action.
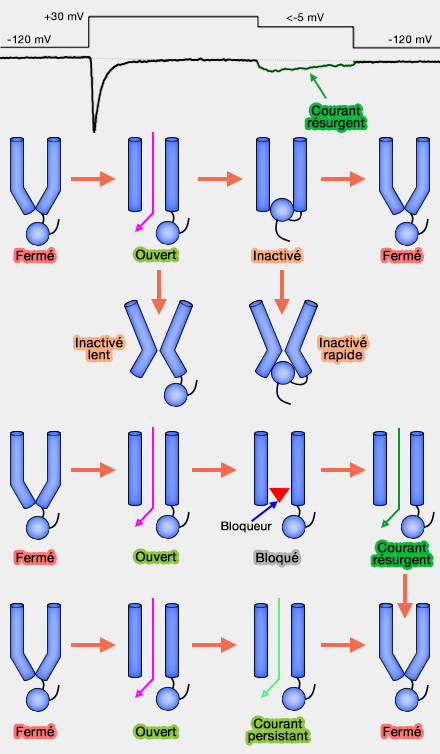
3. Après le déclenchement du potentiel d'action, le canal se désactive rapidement, de l'ordre de la millisecone, en fermant sa porte d'inactivation.
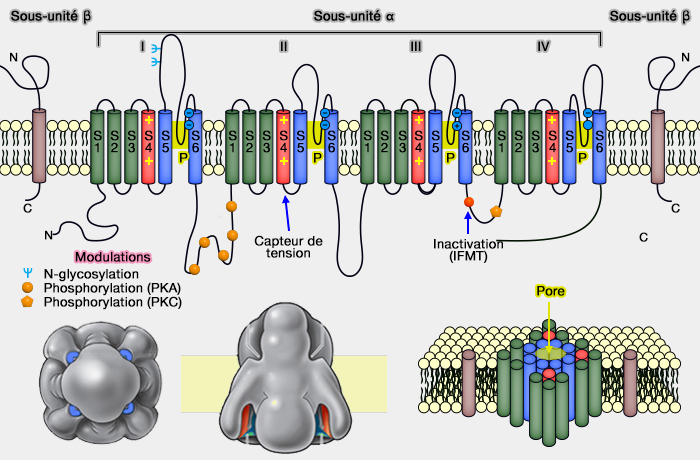
La porte d'inactivation peut être considéré comme une sorte de " bouchon " lié aux domaines III et IV de la sous-unité alpha intracellulaire de la chaîne.
Activation/désactivation des Nav1.7 e t6
(Figure : vetopsy.fr)
Cette fermeture empêchent l'entrée d'autres ions, ce qui provoque l'arrêt de la hausse du potentiel membranaire : le canal est dit inactivé.
Cette phase d'inactivation n'est pas encore comprise complètement.
On peut distinguer :
une inactivation rapide de l'ordre de la milliseconde due à la fermeture de la porte d'activation ;
Les mécanismes transcriptionnels sont régulés par :
des facteurs de transcription agissant sur les promoteurs des gènes,
l'épissage alternatif
Les mécanismes post-transcriptionnels sont représentés par :
la phosphorylation (cf. tableau et figure), en particulier par la PKA (Protein Kinase A) et la PKC (Protein Kinase C), qui peut atténuer les courants membranaire ou non selon les isoformes ;



 Sodium channels, the electrogenisome and the electrogenistat: lessons and questions from the clinic 2012).
Sodium channels, the electrogenisome and the electrogenistat: lessons and questions from the clinic 2012).