La majorité des fibres se dirigent vers les noyaux de l'olive
supérieure.
Le complexe olivaire supérieur
(superior olivary complex) est la structure la plus complexe du système auditif
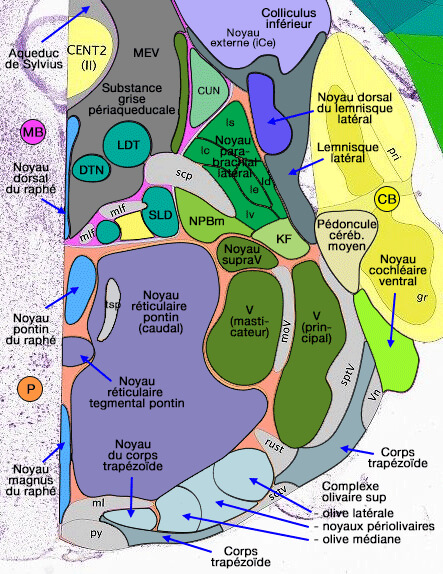
Noyaux du complexe olivaire supérieur
Le complexe olivaire supérieur est composé d'un ensemble de noyaux situés dans la région
ventrale du tegmentum pontique, rostro-ventralement par rapport au noyau du nerf facial (VII).
Le complexe olivaire contient trois noyaux principaux.
1. L'olive latérale supérieure (ou olive principale - LSO -), reconnaissable à sa forme de S, est principalement :
Le corps trapézoïde possède également un noyau ventral et latéral appartenant aux noyaux périolivaires.
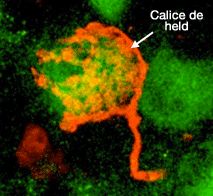
Les cellules globulaires " touffues " (globular bushy cells) contactent les cellules du noyau médian du corps trapézoïde par des synapses géantes, les plus grandes du système nerveux central, appelées calices de Held : elles ressemblent à des calices de fleurs et entourent pratiquement toute la cellule postsynaptique. Le traitement de l'information est dix fois plus rapide que dans la plupart des autres circuits neuronaux.
Ses cellules projettent sur l'olive supérieure latérale ipsilatérale pour l'inhiber grâce à la glycine.
Elles sont à la base du seuil de détection interaural, indispensable à la localisation des sons de hautes fréquences.
Il est à remarquer que ce noyau, réduit chez les primates, est très développé chez les Chauves-souris et les Cétacés.
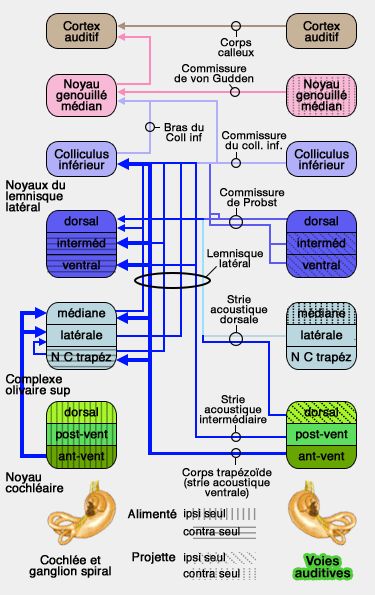
Voies auditives secondaires chez la souris (104/132)
(Figure : vetopsy.fr d'après braininfo.org)
Chez le rat, un quatrième noyau, le noyau paraolivaire supérieur (SPON) ou périolivaire dorso-médial (DMPO), GABAergique et glycinergique, est situé dorsalement dorsal au MNTB.
Le complexe olivaire supérieur est la structure la plus complexe du système auditif.
1. Les noyaux du complexe olivaire supérieur, recevant des afférences des noyaux cochléaires
ventraux, sont impliqués dans l'analyse, la filtration et la modulation de l'information auditive.
Les cellules binaurales (c'est-à-dire répondant
à la stimulation des deux cochlées) sont en charge des caractéristiques
spatiales, c'est-à-dire de la localisation des sons.
Cette analyse s'effectue grâce aux décalages
temporels pour les fréquences basses (inférieures à
3 kHz dans l'olive médiane supérieure ) et d'intensité pour les fréquences hautes (dans l'olive latérale supérieure).
2. Ces noyaux, pour une modulation et une protection des récepteurs auditifs en contrôlant l'entrée du signal sonore dans le circuit neuronal, projettent sur :
Ces deux réflexes, augmentant la rigidité du système tympano-ossiculaire en immobilisant la chaîne des osselets, protègent le tympan en diminuant la pression exercée sur celui-ci
lors de bruits violents (cf. article).




 superior olivary complex) est la structure la plus complexe du système auditif
superior olivary complex) est la structure la plus complexe du système auditif