1. Le NTCP est exclusivement exprimé dans le foie et joue un rôle central dans la circulation entérohépatique des sels biliaires en tant que principal transporteur d'absorption des sels biliaires conjugués du sang (porte) vers le foie.
Les acides biliaires conjugués sont transportés à la fois par Oatp et Ntcp chez la souris, contrairement aux humains où le NTCP est le système dominant d'absorption des acides biliaires.
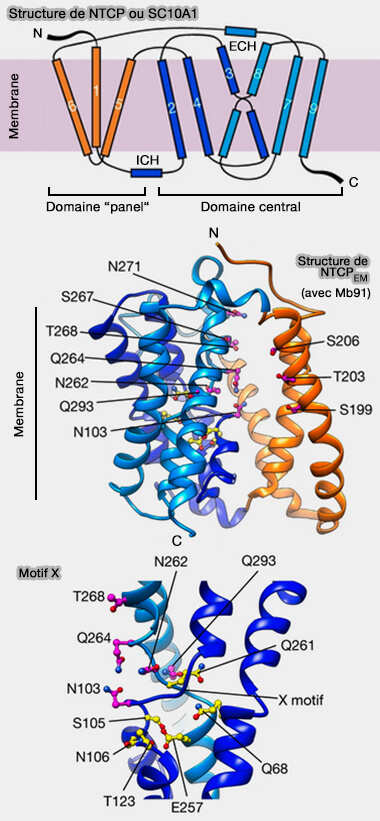
Structure de NTCP
(Figure : vetopsy.fr d'après Goutam et coll)
Les hélices transmembranaires sont reliées par de courtes boucles, ainsi que des hélices α extracellulaires (ECH) et des hélices α intracellulaires (ICH) situées presque parallèlement à la membrane.
1. NTCP est formé par deux domaines structurellement distincts.
a. Le domaine panel ou panneau est formé par TM1, TM5 et TM6 et a perdu la symétrie pseudo-interne par rapport à son équivalent chez les homologues procaryotes SLC10, en raison de la perte évolutive d'une hélice transmembranaire.
b. Le domaine central ou coeur est formé par le regroupement de deux faisceaux d'hélices, TM2–4 et TM7–9.
TM3 et TM8 se déroulent près du milieu de la membrane et se tassent l'un contre l'autre pour former une structure caractéristique en forme de X qui affiche des motifs de résidus polaires hautement conservés parmi les transporteurs de sels biliaires SLC10 des vertébrés.
2. La plupart des résidus rapportés importants pour la liaison du sodium et du substrat correspondent au domaine central.
Les sites de liaison au sodium 1 (Na1, y compris les chaînes latérales S105, N106, T123 et E257) et 2 (Na2, y compris Q68 et Q261), qui ont été observés pour la première fois dans des études cristallographiques d'homologues procaryotes SLC1027, sont structurellement conservés.
La conservation structurale et la mutagenèse NTCP suggèrent fortement que les deux ions sodium, couplés thermodynamiquement au transport des sels biliaires, se lient à ces sites.
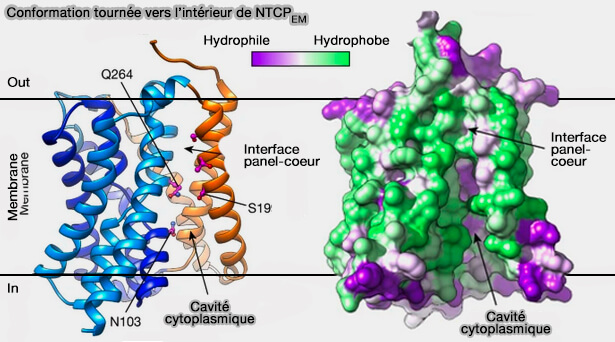
État tourné vers l'intérieur (inward-facing
Dans l'état tourné vers l'intérieur (inward-facing), i.e. en complexe avec Nb87, les domaines de noyau et de panneau de NTCPEM sont étroitement serrés les uns contre les autres du côté extracellulaire de la membrane.
Du côté intracellulaire, les domaines se séparent, découvrant une grande cavité amphiphile, i.e. 1 500 Å3, qui s'ouvre sur le cytoplasme, ainsi que latéralement sur le noyau hydrophobe de la membrane à travers une crevasse entre TM6 et TM9.
De l'autre côté du transporteur, TM1 et TM5 se tassent contre le domaine central, obstruant la cavité de la membrane.
Conformation tournée vers l'intérieur (inward-facing) de NTCPEM
(Figure : vetopsy.fr d'après Goutam et coll)
La conservation des acides aminés, les études de mutagenèse et le grand volume de la cavité suggèrent qu'elle fait partie de la voie du substrat du côté cytoplasmique.
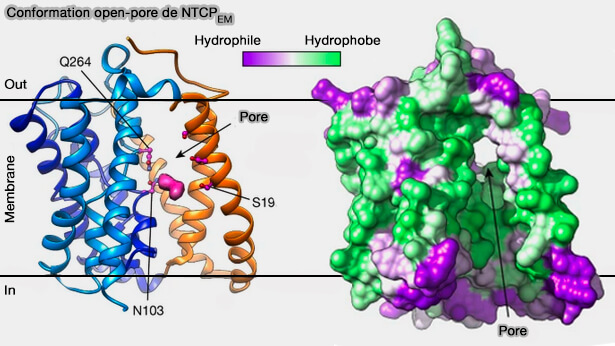
Dans la conformation pore-ouvert (open-pore), i.e. en complexe avec Mb91, NTCPEM montre un changement conformationnel marqué par rapport à l'état orienté vers l'intérieur
Les domaines du noyau et du panneau tournent autour de 20º et se déplacent d'environ 5 Å vers les côtés opposés de la membrane sous forme de corps presque rigides.
Ces mouvements sont facilités par les résidus glycine et proline conservés qui agissent comme des charnières dans les boucles de connexion, ainsi que dans l'ICH et l'ECH .
1. En conséquence, les deux domaines se séparent des côtés extracellulaire et cytoplasmique et ouvrent un large pore à travers le transporteur, exposant les sites de liaison Na+ et les résidus de motif X simultanément aux côtés opposés de la membrane.
Il s'agit d'une transition conformationnelle inattendue, car les transporteurs actifs alternent généralement l'exposition de leurs sites de liaison de ligand aux milieux extracellulaire et intracellulaire et adoptent des états intermédiaires avec des substrats occlus dans la protéine.
Conformation pore-ouvert (open-pore) de NTCPEM
(Figure : vetopsy.fr d'après Goutam et coll)
2. Le pore a un diamètre minimum d'environ 5 Å et contient un grand volume, i.e. 2 400 Å3) avec son axe long orienté à un angle d'environ 45º par rapport au plan de la membrane.
La surface tapissant le pore est amphiphile et la plupart des résidus polaires de cette surface proviennent du domaine central, y compris les chaînes latérales conservées dans le motif X.
La conservation des acides aminés, l'architecture et la nature amphiphile du pore suggèrent fortement qu'il s'agit de la voie de translocation d'un large éventail de substrats volumineux amphiphiles transportés par le NTCP.
Les NTCP transportent outre les sels biliaires, les stéroïdes sulfatés et les statines.
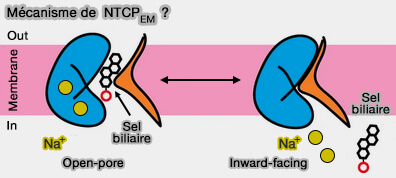
Mécanisme ?
La structure à pores ouverts du NTCPEM est apparemment en contradiction avec les mécanismes de transport à accès alterné observés dans la plupart des familles de porteurs de solutés qui impliquent des intermédiaires liés au substrat occlus du cycle de transport.
Mécanisme de NTCP ?
(Figure : vetopsy.fr d'après Goutam et coll)
Comment concilier un état intermédiaire à pores ouverts avec un transport thermodynamiquement actif ?
Le pore pourrait être ouvert de manière transitoire en présence de substrat et de Na+ couplé thermodynamiquement.
Le pore se fermerait lors de la libération de ligands dans le cytoplasme à l'état orienté vers l'intérieur.
En outre, les ions sodium contribueraient à fermer le pore, évitant la perméation des sels biliaires dans son gradient électrochimique.
Acides biliaires dans le foie
Dans le foie, les acides biliaires libres sont reconjugués et les acides biliaires, i.e. réabsorbés et nouvellement synthétisés, sont sécrétés dans l'intestin avec la bile.
De petites quantités d'acides biliaires ne pas captées par les hépatocytes et entrent dans la circulation sanguine par la veine hépatique, i.e. les acides biliaires sont aussi présents :
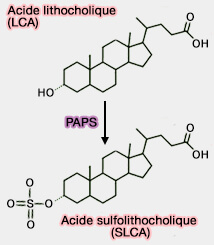
Sulfoconjugaison de
l'acide lithocholique
(Figure : vetopsy.fr)
dans le sang, les urines, les selles, dans lesquels ils peuvent servir aux analyses,
le liquide amniotique, les expectorations, les épanchements pleuraux et le liquide de lavage bronchoalvéolaire.
Excrétion des acides biliaires dans les selles
Une partie des acides biliaires primaires et secondaires déconjugués est également excrétée dans les fèces (environ 600 mg/24h).