Bien que possédant un capteur de tension, les canaux CNG ne sont pas bloqués par la tension, une propriété essentielle pour une phototransduction et une transduction olfactive appropriées.
Le canal CNG n'est pas voltage-dépendant pour deux raisons.
1. Dans un VGIC, le modèle classique est le modèle d'hélice de glissement (" sliding helix ") ou vis hélicoïdale (" helical screw ") qui nécessite la formation séquentielle de paires ioniques ( structure et fonctionnement de S4).
Cette structure hélicoïdale est continue et forme un angle de 65° avec la membrane.
Le S4 du CNG se divise en trois sous-segments, S4a (boucle), S4b (hélice 310) et S4c (hélice α) qui possèdent des orientations différentes : certaines charges positives sont ainsi " occultées ", tandis que d'autres sont engagés dans des interactions avec S2, ce qui, au global, ne leur permettent pas de détecter la tension membranaire, (Structure of a eukaryotic cyclic nucleotide-gated channel 2017).
2. Le lien S4-S5 (S4-S5 linker) typique dans les VGIC est une hélice α de 10 à 12 acides aminés parallèle au plan de la membrane.
Le lien S4-S5 interagit avec S6, une interaction cruciale pour le déclenchement dépendant de la tension (rôles du lien S4-S5).
Le lien S4-S5 des CNG, composé que de 5 acides aminés, n'a aucune interaction avec S6, mais forme une boucle courte. S4, S5 et le lien S4-S5 adoptent un motif hélice-coude-hélice qui entre en contact avec le lien C, i.e. le VSLD semble réellement impliqué dans la synchronisation des ligands des canaux CNG.
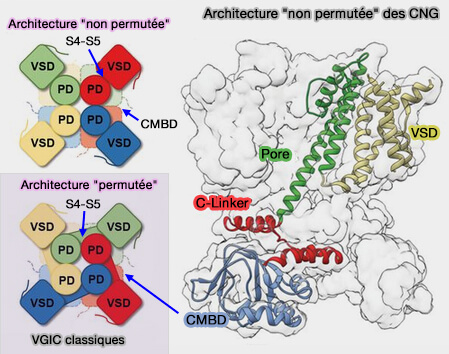
Liaison VSD-pore non " permutée "
(Figure : vetopsy.fr d'après James et coll)
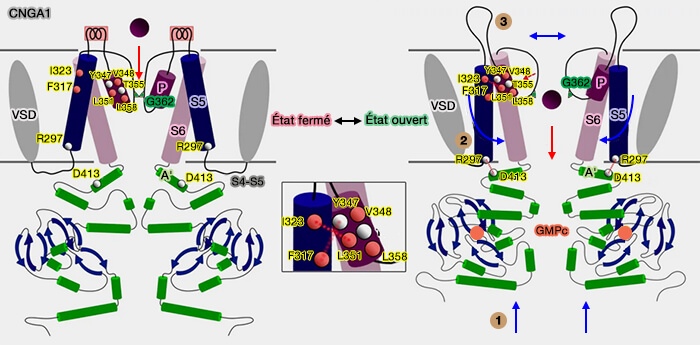
Le déclenchement des canaux CNG est initié par la liaison du nucléotide cyclique à son domaine de liaison (CNBD) et se termine par l'ouverture de la porte située dans la région des pores, à environ quelques dizaines d'Å de distance.
1. Lorsque le GMPc se délie, i.e, lorsque le canal a accueilli son ligand et le pore de sélectivité est ouvert, l'hélice C du CNBD se déplace vers le bas (figure 1).
2. Les hélices E'F' dans le lien-C se déplacent vers le bas, entraînant des changements conformationnels dans les hélices C'D' dont le résultat est un changement de conformation de S6 par deux modèles possibles :
soit les interactions entre C'D 'et A'B' sont affaiblies, provoquant un affaiblissement des interactions entre A'B' et le TMD (figure 2) ;
soit C′D′ et A′B ′ se dissocient, provoquant la dissociation de A′B′ du TMD (figure 3).
Modèle de l'ouverture des canaux CNG
(Figure : vetopsy d'après Li and coll)
3. Les hélices A'B'C'D' constituent le gating ring qui modifie sa conformation et ses interactions avec le domaine transmembranaire (TMD) lors de la liaison ou du détachement des nucléotides cycliques. Ces changements de conformation sont transmis directement à S6.
4. Les mouvements de S6 perturbent les interactions hydrophobes entre S6, l'hélice des pores et le filtre de sélectivité, ce qui provoque sa fermeture.
Deuxième modèle
Un deuxième modèle implique que le signal transmis du CNBD au gating ring situé dans la région du filtre de sélectivité implique des changements conformationnels de S5, des changements conformationnels de S5, et non une flexion vers l'extérieur de la partie intracellulaire de S6. Cette étude a été réalisé sur le canal CNGA1, en mutant certains résidus (The gating mechanism in cyclic nucleotide-gated ion channels 2018).
Pendant le déclenchement, il y a des changements conformationnels de S5, dans les boucles extracellulaires reliant S5 à l'hélice P et dans les boucles intracellulaires reliant S4 à S5. La comparaison se fait avec le TAX-4 décrit dans le premier modèle et tiré de Structure of a eukaryotic cyclic nucleotide-gated channel (2017).
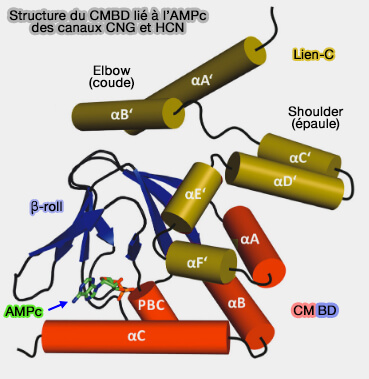
Hélice A' du Lien-C de CNGA1
(Figure : vetopsy.fr d'après Mazzolini et coll)
Le résidu R308 de TAX-4 (équivalent au R297 de CNGA1) dans l'hélice S5 et D429 (équivalent à D413) dans l'hélice A' sont impliqués dans un pont salin, couplant les changements de conformation dans le CNBD à la synchronisation des canaux comme ce qui a été observé dans les canaux HCN.
1. La liaison des nucléotides cycliques est suivie de réarrangements dans le lien-C : les hélices A' des sous-unités voisines s'écartent les unes des autres.
2. Le mouvement des résidus D413 dans les hélices A’ vers R297 dans S5 favorise les interactions électrostatiques entre ces résidus.
3. Ces interactions électrostatiques induisent des changements de conformation principalement dans S5.
Changement de flexibilité de S5 de CNGA1
(Figure : vetopsy.fr d'après Mazzolini et coll)
4. Ces changements de conformation sont transmis à l'hélice P par des interactions hydrophobes entre S5 (I309, H310, Y316, F317) et l'hélice P (L343, Y347, L351, L356 et L358).
5. Les changements conformationnels dans l'hélice P, suite à la la liaison du GMPc, élargissent la lumière des pores.





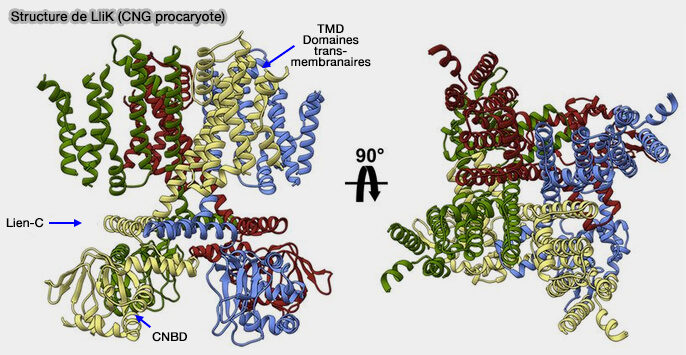
 fonctionnement du capteur de tension et pore et son fonctionnement).
fonctionnement du capteur de tension et pore et son fonctionnement).