


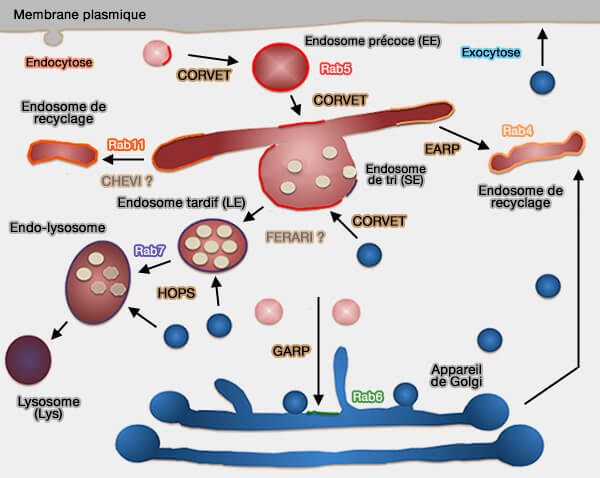
Système endo-lysosomal
Endosomes précoces
Complexes de tri ou de recyclage :
CHEVI et FERARI
- Vue d'ensemble du système endomembranaire
- Endosomes
- Vue d'ensemble des endosomes
- Endosomes précoces
- Vue d'ensemble des endosomes précoces
- Devenirs des endosomes précoces
- Composants des endosomes précoces
- Rab5 et ses régulateurs
- Rab5 et ses effecteurs
- Complexes d'attache des endosomes précoces : complexe CORVET
- Complexe de tri et de recyclage des endosomes précoces
- Endosomes de recyclage
- Vésicules intraluminales (ILV) et endosomes ou corps mutivésiculaires (MVE/MVB)
- Endosomes tardifs
- Système endo-lysosomal et phosphoinositides
- MCS (sites de contact membranaire) RE/endosomes/lysosomes
- Lysosomes
- Trafic des endosomes/lysosomes
- Fusion et fission membranaire
- Transport membranaire
- Moteurs moléculaires
- Voies de signalisation

Des complexes de tri ou de recyclage des endosomes précoces, autres que CORVET et HOPS, i.e. CHEVI ET FERARI, peuvent différer par leur module d'interaction Rab ou les protéine SM.
Les complexes les plus connus dans le système endo-lysosomal sont :
- le complexe CORVET des endosomes précoces,
- le complexe HOPS des endosomes tardifs.
Toutefois, d'autres complexes peuvent différer par leur module d'interaction Rab ou la protéine SM.
Par exemple, chez la levure, CORVET et HOPS pourraient être interconvertis par le biais de complexes mixtes intermédiaires (The CORVET Tethering Complex Interacts with the Yeast Rab5 Homolog Vps21 and Is Involved in Endo-Lysosomal Biogenesis 2007).
(Figure : vetopsy.fr d'après Spang)
Complexe CHEVI (class C Homologs in Endosome-Vesicle Interaction)
Structure de CHEVI
1. Chez les métazoaires, on trouve deux homologues de Vps16 :
- Vps16,
- Vps16B, appelé aussi SPE39, SPE-39, VIPAR, VPS33B interacting protein, apical-basolateral polarity regulator.
2. Le consensus actuel est que Vps16 est présent à la fois dans les complexes CORVET et HOPS (Characterization of the Mammalian CORVET and HOPS Complexes and Their Modular Restructuring for Endosome Specificity 2015 et Recruitment of VPS33A to HOPS by VPS16 Is Required for Lysosome Fusion with Endosomes and Autophagosomes 2015).
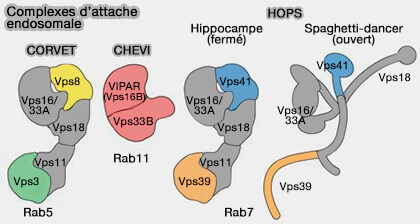
(Figure : vetopsy.fr d'après van der Beck et coll)
3. Le complexe CHEVI est formé par Vps16B avec Vps33B.
- Vps16B/SPE-39 a d'abord été découverte chez Caenorhabditis elegans : cette protéine se lie à Vps33 et cette interaction est essentielle au trafic vésiculaire au cours de la spermatogenèse (The Caenorhabditis elegans spe-39 Gene Is Required for Intracellular Membrane Reorganization During Spermatogenesis 2003).
- L'orthologue humain de SPE-39 (hSPE-39) est également connu pour s'associer aux deux sous-types de Vps33A et Vps33B dans les cellules de mammifères (SPE-39 Family Proteins Interact with the HOPS Complex and Function in Lysosomal Delivery 2009 et Metazoan cell biology of the HOPS tethering complex 2011 et Inhibitory effect of SPE-39 due to tyrosine phosphorylation and ubiquitination on the function of Vps33B in the EGF-stimulated cells 2012).

Il semble que CHEVI ne contienne que ces deux unités, mais il se peut que nous n'ayons pas trouvé les autres.
2. Des échanges entre les protéines SM, i.e. protéines de la famille ou Sec1/Munc18 comme Vps33A et B, pourraient se produire dans le complexe d'attache.
Cette notion serait cohérente avec les observations chez C. elegans et pourrait expliquer comment CORVET peut affecter différentes étapes d'attache dans les cellules de mammifères (Mammalian CORVET Is Required for Fusion and Conversion of Distinct Early Endosome Subpopulations 2014).
Rôle de CHEVI dans le recyclage
CHEVI est localisé dans les cellules polarisées.
1. Par contre, on ne sait toujours pas dans quelle étape de recyclage CHEVI est impliquée, c'est-à-dire s'il s'agit d'endosomes précoces ou d'endosomes de recyclage vers la membrane plasmique, ou les deux.
- VPS33B et Vps16B se localiseraient au niveau des endosomes de recyclage RAB11+ (Associations Among Genotype, Clinical Phenotype, and Intracellular Localization of Trafficking Proteins in ARC Syndrome 2012).
- Leur KO altère la localisation correcte des protéines apicales, entraînant une polarisation cellulaire anormale (Vps33b is crucial for structural and functional hepatocyte polarity 2017).
RAB11 interagit à la fois avec Vps33A et Vps16B, mais ne les recrute dans les membranes que lorsqu'elles sont assemblées dans le complexe CHEVI (Mutations in VIPAR cause an arthrogryposis, renal dysfunction and cholestasis syndrome phenotype with defects in epithelial polarization 2010).
2. Le complexe CHEVI ne se localise pas significativement dans les endosomes tardifs, mais il peut quand même être recruté par RILP (RAB7 interacting lysosomal protein) du complexe HOPS des mammifères dans les lysosomes (Late endosomal transport and tethering are coupled processes controlled by RILP and the cholesterol sensor ORP1L 2013 et Characterization of the Mammalian CORVET and HOPS Complexes and Their Modular Restructuring for Endosome Specificity 2015).
La surexpression de RILP induit le recrutement de Vps33B dans les endosomes tardifs et les lysosomes (Vps33B is required for delivery of endocytosed cargo to lysosomes 2015).
- Comme les endosomes de recyclage dédiés à Rab11 positifs ne se trouvent que dans un sous-ensemble de types cellulaires, cela implique que la localisation de Vps33B diffère selon les types cellulaires (The Complex Ultrastructure of the Endolysosomal System 2014).
- En outre, il semble que l'expression et la localisation de Vps33B dépendent du statut métabolique de la cellule, ouvrant la possibilité que la fonction et la localisation de Vps33B soient sensibles au statut nutritionnel.
3. Le complexe CCC et le complexe retriever sont impliqués dans le tri des cargos portant un motif NPxY/NxxY, comme les intégrines, dans une voie de recyclage endosomale spécialisée.
Vps33B :
- interagit avec CCDC22 du complexe CCC (Proteomic and Biochemical Comparison of the Cellular Interaction Partners of Human VPS33A and VPS33B 2018),
- lie les sous-unités de l'intégrine (Characterization of a Novel Integrin Binding Protein, VPS33B, Which Is Important for Platelet Activation and In Vivo Thrombosis and Hemostasis 2015),
- est nécessaire au recyclage de l'intégrine, comme dans les cellules épidermiques granulaires de la peau dans lesquelles le complexe CHEVI est requis pour une biogenèse et/ou une sécrétion correctes des corps lamellaires, essentiels à la fonction de barrière cutanée, et ceci en interaction avec Rab11 (VPS33B and VIPAR are essential for epidermal lamellar body biogenesis and function 2018).
Vps33B a-t-il la capacité de recruter le complexe CCC dans les endosomes de recyclage Rab11-positifs pour la formation de tubules de recyclage ?
Rôles dans la biogenèse des LRO
CHEVI est requis pour la biogenèse de certains types d'organites liés aux lysosomes, appelés LRO ou Lysosome-Related Organelles (The CHEVI tethering complex: facilitating special deliveries 2016 pour une revue).
1. Dans les mégacaryocytes, i.e. cellules progénitrices des plaquettes (thrombocytes) qui contiennent deux types de LRO, les granules denses et les granules α, le complexe CHEVI est requis pour :
- la livraison du contenu des granules aux compartiments endosomaux agissant comme précurseurs des granulés α, i.e. transport des protéines de cargaison du réseau trans-Golgi (TGN) vers les organites progéniteurs des granules α,
- de ces compartiments aux granulés α eux-mêmes,
- pour le transport des granules α dans les extensions proplaquettaires.
Il existerait aussi des interactions avec la α-tubuline et SEC22B (VPS33B regulates protein sorting into and maturation of α-granule progenitor organelles in mouse megakaryocytes 2015).
Des mutations autosomiques récessives dans VPS33B ou VPS16B provoquent le syndrome ARC (Syndrome d'arthrogrypose insuffisance rénale cholestase).
- L'ARC est un trouble multisystémique mortel (dysfonctionnement rénal et cholestase) caractérisé par des défauts de transport apical dans les cellules polarisées (Mutations in VPS33B, encoding a regulator of SNARE-dependent membrane fusion, cause arthrogryposis–renal dysfunction–cholestasis (ARC) syndrome 2004).
- Le phénotype résultant est similaire au syndrome des plaquettes grises (GPS ou Gray Platelet Syndrome), le trouble plaquettaire héréditaire classique causé par l'absence de granules.

Vous pouvez lire : CORVET, CHEVI and HOPS – multisubunit tethers of the endo-lysosomal system in health and disease (2019).
2. Dans les cellules des canaux collecteurs du rein, CHEVI est nécessaire pour le transport de l'enzyme LH3 (Lysyl Hydrolase 3) modifiant le collagène, appelée aussi PLOD3, vers des compartiments spécialisés qui contiennent également du collagène IV (Regulation of post-Golgi LH3 trafficking is essential for collagen homeostasis 2016).
Ces compartiments présentent des caractéristiques des LRO, bien que leur mode de biogenèse soit encore obscur.
CHEVI assure la médiation de deux étapes de trafic de LH3 :
- sortie dépendante de Rab10 du réseau trans-Golgi (TGN),
- étape de tri supplémentaire dépendante de Rab25, potentiellement des endosomes vers les compartiments du collagène IV (Characterization of the Role Rab25 in Energy Metabolism and Cancer Using Extracellular Flux Analysis and Material Balance 2015).
La perturbation du complexe CHEVI dans ces cellules entraîne une perturbation de la matrice extracellulaire (ECM) et une polarisation cellulaire aberrante.
Complexe FERARI
Les protéines SM, i.e. protéines de la famille ou Sec1/Munc18, contrairement à celles de CORVET et HOPS, ne sont pas un composant intrinsèque de la plupart des autres attaches.
Par conséquent, il est possible qu'il existe encore d'autres types de complexes qui portent Vps45, la troisième protéine SM du système endosomal.
(Figure : vetopsy.fr d'après Spang)
1. FERARI (Factors for Endosome Recycling And Retromer Interactions) pourrait être un autre complexe contenant Vps16B et Vac1/rabenosyne-5 qui ont été identifiés comme des partenaires de liaison non SNARE de Vps45 (The SM protein VPS-45 is required forRAB-5-dependent endocytic transport in Caenorhabditis elegans 2007).
- Ce complexe d'attache pourrait représenter un complexe de type HOPS et CORVET minimal, car Vac1/rabenosyne-5 médie l'interaction entre Vps21/Rab5 et PI(3)P (Rabenosyn-5, a Novel Rab5 Effector, Is Complexed with hVPS45 and Recruited to Endosomes through a FYVE Finger Doma 2000).
- Comme HOPS et CORVET, ce complexe contient un module d'interaction Rab et un complexe SNARE.
2. Ce complexe pourrait agir sur les endosomes en favorisant le recyclage et le transport rétrograde ou centripète vers le réseau trans-Golgi (TGN) d'une manière rétromère-dépendante (Common and distinct roles for the binding partners Rabenosyn-5 and Vps45 in the regulation of endocytic trafficking in mammalian cells 2010).
Retour aux endosomes précoces
Biologie cellulaire et moléculaireMembrane plasmiqueNoyauCytoplasmeMitochondriesSystème endomembranaireRéticulum endoplasmiqueAppareil de GolgiEndosomesLysosomesPeroxysomesProtéasomesCytosquelettemicrofilaments d'actineFilaments intermédiairesMicrotubulesMatrice extracellulaireReproduction cellulaireBiochimieTransport membranaire Moteurs moléculairesVoies de signalisation