


Système endo-lysosomal : endosomes précoces
Complexes de tri ou de recyclage :
interactions du rétromère
- Vue d'ensemble du système endomembranaire
- Endosomes
- Vue d'ensemble des endosomes
- Endosomes précoces
- Vue d'ensemble des endosomes précoces
- Devenirs des endosomes précoces
- Composants des endosomes précoces
- Rab5 et ses régulateurs
- Rab5 et ses effecteurs
- Complexes d'arrimage des endosomes précoces : complexe CORVET
- Complexe de tri et de recyclage des endosomes précoces
- Endosomes de recyclage
- Vésicules intraluminales (ILV) et endosomes ou corps mutivésiculaires (MVE/MVB)
- Endosomes tardifs
- Système endo-lysosomal et phosphoinositides
- MCS (sites de contact membranaire) RE/endosomes/lysosomes
- Lysosomes
- Trafic des endosomes/lysosomes
- Fusion et fission membranaire
- Transport membranaire
- Moteurs moléculaires
- Voies de signalisation

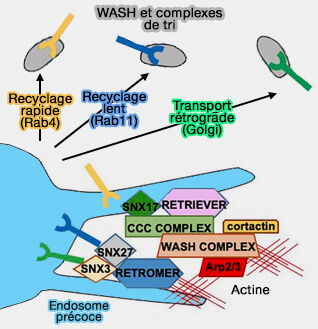
Le rétromère est un complexe de protéines hautement conservé par l'évolution qui joue un rôle important dans le du tri et du trafic des endosomes précoces (Quick guide : retromer 2017).
Vue d'ensemble du recrutement du CSC
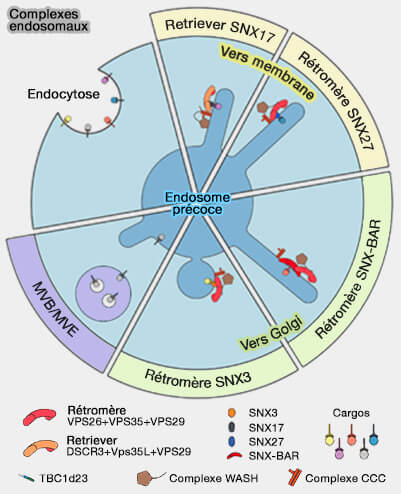
Le rétromère est recruté sur les endosomes par des motifs d'acides aminés courts ou de signaux de tri trouvés dans les protéines membranaires.
(Figure : vetopsy.fr d'après Wang et all)
Comme le rétromère ne peut se lier directement aux lipides membranaires, il doit utiliser de multiples interactions protéine/protéine.
1. Des adaptateurs de cargos peuvent être associés à la membrane.
2. Le rétromère peut utiliser des protéines accessoires comme (A mechanism for retromer endosomal coat complex assembly with cargo 2014) :
- les nexines (SNX),
- les petites GTPases Rab,
- le complexe WASH.
Le rétromère interagit aussi avec Varp, protéine se liant à la golginA4, au moteur kinésine Kif5a et à Vps29 (A Molecular Network for the Transport of the TI-VAMP/VAMP7 Vesicles from Cell Center to Periphery 2012).
Remarque : la liaison aux cargos s'effectue en général par VPS35, mais aussi par VPS26, comme dans le cas du transporteur DMT1-II (Divalent Metal Transporter1) couplé à SNX3 ( rétromère/SNX3)
rétromère/SNX3)
Nexines liées au rétromère
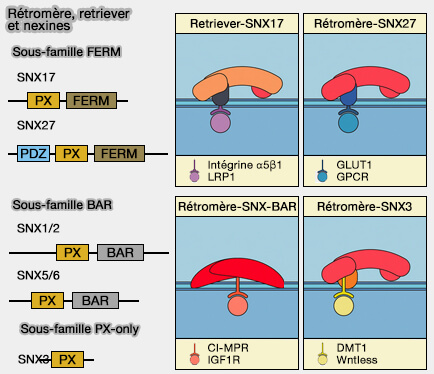
Les nexines (SNX) sont des protéines associées à la membrane, qu'on peut considérer comme des protéines adaptatrices à domaine PX (PhoX homology).
Plusieurs nexines s'attachent au rétromère (Unveiling the cryo-EM structure of retromer 2020) pour établir une spécificité du cargo, du tri, du remodelage membranaire et du trafic.

Les nexines sont traitées dans un chapitre spécial.
(Figure : vetopsy.fr d'après Wang et all)
1. Le rétromère canonique (rétromère/SNX-BAR) est attaché à un dimère de nexine appartenant à la sous-famille BAR :
- SNX1 ou SNX2, i.e. équivalent à Vps5 de la levure,
- SNX5, SNX6, i.e. équivalent à Vps17 de la levure.
2. Le rétromère/SNX3 est formé par le rétromère lié à SNX3, une nexine contenant seulement un domaine PX (sous-famille PX-only), qui, quoique sans domaine BAR, peut provoquer une tubulation in vitro.

Ces deux voies de transport rétrograde conduit les cargos au réseau trans-Golgi (TGN) via le réseau de microtubules (Microtubule motors mediate endosomal sorting by maintaining functional domain organization 2013).
3. Le rétromère/SNX27 est formé par le rétromère lié à SNX27, une nexine appartenant à la sous-famille FERM.
Cette voie recycle les cargos directement vers la membrane plasmique via le réseau de microtubules.
Remarque : Snx17 est impliqué dans le complexe retriever, un complexe rétromère-like.

La complexité de la biologie des rétromères chez les métazoaires reflète probablement à la fois l'augmentation du nombre de cargos et la diversité des routes de trafic à partir des endosomes.
Petites GTPases Rab
Les petites GTPases, essentielles à la formation de vésicules endocytaires enveloppées, aussi bien à base de clathrine, de COPI et COPII, sont aussi des régulateurs du trafic vésiculaire médié par le rétromère.
Rab7 et TBC1D5
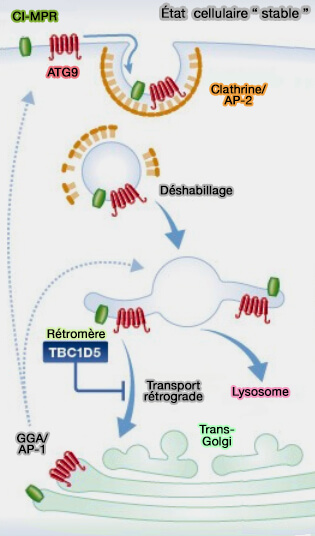
La petite GTPase Rab7a endosomal est requise pour le recrutement membranaire du rétromère canonique (rétromère/SNX-BAR) chez la levure, les plantes et les mammifères.
(Figure : vetopsy.fr d'après Popovic et Dikic)
1. TBC1d5, membre de la famille TBC (Tre2-Bub2-Cdc16) qui comprend plus de 40 membres chez l'homme, est un GAP (GTPase-activating protein) de Rab7a, i.e. catalysant l'hydrolyse du GTP et donc inactivant Rab7a (Structural and mechanistic insights into regulation of the retromer coat by TBC1d5 2016).
La surexpression de TBC1d5 diminue la quantité de Rab7a-GTP et réduit la quantité de VPS35 sur les endosomes (Membrane recruitment of the cargo-selective retromer subcomplex is catalysed by the small GTPase Rab7 and inhibited by the Rab-GAP TBC1D5 2009).
2. De tous les régulateurs endogènes connus du rétromère, TBC1d5 affiche la plus grande affinité envers le rétromère (constante de dissociation de 200 nM à 450nM), i.e. comparable à celle mesurée entre VPS29 et VPS35 (200 nM), et est supérieure d'au moins un ordre de grandeur à l'affinité entre le rétromère et ses autres partenaires de liaison, tels que SNX3, VARP, Rab7a ou la sous-unité du complexe WASH, FAM21.
TBC1d5 abrite un domaine TBC sur son extrémité N-terminale et une extrémité C-terminale largement désordonnée (Specific TBC Domain-Containing Proteins Control the ER-Golgi-Plasma Membrane Trafficking of GPCRs 2019).
Le domaine TBC assure la médiation de l'interaction avec le rétromère, en contactant à la fois VPS35 et VPS2921.
- Une boucle de TBC1d5 se lie à une poche hydrophobe conservée sur VPS29, et une seconde boucle peut interagir avec l'extrémité N-terminale de VPS35.
- Un mutant VPS35 incapable de s'associer à TBC1d5 ne s'est pas correctement localisé dans les endosomes, ce qui suggère que TBC1d5 pourrait jouer un rôle dans le recrutement de VPS35 dans la membrane endosomale.
L'association étroite entre TBC1d5 et le rétromère s'apparente au complexe Sec23-Sec24 dans le manteau COPII.
La plupart des protéines TBC utilisent un mécanisme catalytique à double doigt Arg-Gln pour inactiver les substrats Rab qui est largement différent du mécanisme à doigt Arg unique utilisé par de nombreux GAP des autres petits GTPases.
Remarque : Le domaine TBC se retrouve dans d'autres TBC comme TBC1d6, qui module l'interaction de Rab26 avec le trafic post-Golgi des récepteurs couplés à une protéine G ou GPCR (Specific TBC Domain-Containing Proteins Control the ER-Golgi-Plasma Membrane Trafficking of GPCRs 2019).
(Figure : vetopsy.fr d'après Popovic et Dikic)
3. Le rétromère et TBC1d5 pourrait avoir un rôle dans la régulation de l'activation et de la localisation subcellulaire de Rab7a (Control of RAB7activity and localization throughthe retromer-TBC1D5complex enablesRAB7-dependent mitophagy 2017).
Rab7a est localisé sur plusieurs organites :
- les endosomes tardifs/lysosomes LAMP1/2+,
- les endosomes de tri EEA1+,
- le réseau Trans-golgi (TGN),
- le réticulum endoplasmique (RE),
- les mitochondries.
En l'absence de rétromère ou de TBC1d5, les niveaux de Rab7a-GTP augmentent considérablement et Rab7a s'accumule sur les domaines lysosomaux, ce qui entraîne une diminution de la mobilisation de Rab7a, un épuisement de Rab7a inactif des autres organites subcellulaires et un renouvellement membranaire défectueux.
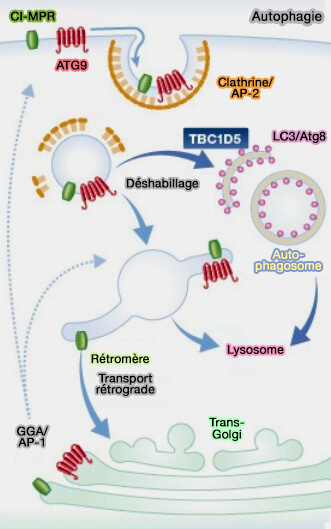
4. TBC1d5 joue également un rôle dans l'autophagie.
- TBC1d5 oscille entre les endosomes et les autophagosomes (TBC1D5 and the AP2 complex regulate ATG9 trafficking and initiation of autophagy 2014).
- L'association TBC1d5 avec les autophagosomes favorise le recyclage de GLUT1 et l'absorption de glucose (Autophagy-Dependent Shuttling of TBC1D5 Controls Plasma Membrane Translocation of GLUT1 and Glucose Uptake 2017).
Le TBC1D15 endogène se localise principalement dans les membranes mitochondriales, où il participe à la mitophagie et à la fission mitochondriale organisée par le réticulum endoplasmique (Mitochondrial Rab GAPs govern autophagosome biogenesis during mitophagy 2014 et Mitochondria-lysosome contacts regulate mitochondrial fission via Rab7 GTP hydrolysis 2018).
Le TBC1D15 mitochondrial semble coexister en équilibre avec son pool cytosolique et Arl8b/SKIP/HOPS puise dans ce dernier pour procéder à l'élimination de Rab7 de certains MVB.
Autres Rab
1. Rab21 est impliqué dans le tri des cargos en formant un complexe avec WASH et le rétromère pour réguler l'actine F endosomale (APEX2‐mediated RAB proximity labeling identifies a role for RAB21 in clathrin‐independent cargo sorting 2019).
VARP (Vps9- ankyrin repeat protein), protéine cytosolique de 1050 acides aminés, s'est avéré être (Structure/Function Studies on Vps9 domain-containing Ankyrin Repeat protein (VARP) 2019) :
- un GEF de Rab21,
- un effecteur Rab32/38,
- un inhibiteur de la R-SNARE VAMP7.
2. Rab32/38 sont impliqués dans le trafic du transporteur de glucose GLUT1 vers la membrane plasmique en régulant la molécule effectrice VARP, qui à son tour se lie au v-SNARE TI-VAMP/VAMP7 (Role of Varp, a Rab21 exchange factor and TI-VAMP/VAMP7 partner, in neurite growth 2009), pour faciliter la fusion membranaire entre les endosomes de recyclage portant GLUT1 (VARP Is Recruited on to Endosomes by Direct Interaction with Retromer, Where Together They Function in Export to the Cell Surface 2014)
(Figure : vetopsy.fr d'après Capitani et coll)
Remarque : BLOC-3 est le principal GEF pour Rab32/Rab38 (BLOC-3 et Rab32/38)
3. SNX1 interagirait avec Rab6IP, une protéine interagissant avec Rab6 localisée dans le compartiment de Golgi et impliquée dans l'attachement des transporteurs dérivés d'endosomes au TGN (Rab6-interacting Protein 1 Links Rab6 and Rab11 Function 2007)
Complexe WASH
Le complexe WASH est étudié dans un chapitre spécial.
Fonctions du rétromère
Biologie cellulaire et moléculaireMembrane plasmiqueNoyauCytoplasmeMitochondriesSystème endomembranaireRéticulum endoplasmiqueAppareil de GolgiEndosomesLysosomesPeroxysomesProtéasomesCytosqueletteMicrofilaments d'actineFilaments intermédiairesMicrotubulesMatrice extracellulaireReproduction cellulaireBiochimieTransport membranaireMoteurs moléculairesVoies de signalisation