Dans les transporteurs primitifs, comme dans les SemiSWEET, les faisceaux d'hélice constituant chaque pièce du mécanisme ne bougent pas en tant que corps rigide strict, mais peuvent légèrement se plier.
Comme le résidu proline de la boucle PQ (PQ-loop) est hautement conservé dans les SWEET et les transporteurs de la famille PQ-loop, ce mouvement pourrait être conservé chez tous les ces transporteurs.
Ces mécanismes sont alors asymétriques.
Mécanisme asymétrique simple archaïque
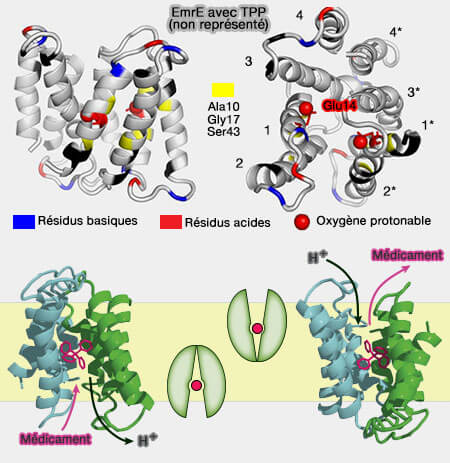
EmrE
Structure et antiport de EmrE
(Figure : vetopsy.fr d'après Morrison et coll)
1. EmrE se compose de quatre segments transmembranaires (TM1 à TM4) qui s'insèrent dans la membrane aussi bien avec leur extrémités N- et C-terminales à l'intérieur qu'à l'extérieur (topologies Nin-Cin et Nout-Cout).
Leur disposition membranaire dimérique est encore sujette à discussion.
Ce modèle d'interrupteur à bascule antiparallèle aurait ainsi la possibilité de positionner des polypeptides asymétriques plus facilement autour du site de liaison au substrat
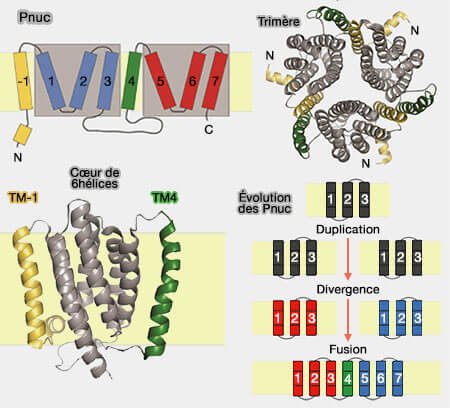
Pnuc
Structure et évolution des Pnuc
(Figure : vetopsy.fr d'après Jaehme et Slotboom)
1. Chaque protomère est constitué de domaines transmembranaires, numérotée de -1 à 7, car le premier segment est absent de nombreux membres des PnuC et de tous les membres de la famille SWEET.
Les TM1 à 3 et TM5 à 7 sont analogues aux TM1 à 3 des SWEET, mais chaque groupe est inversé et présente une homologie faible.
Certains homologues semblent avoir 13, 14, 15 ou 16 TM, certaines perméases MFS sont fusionnées à d'autres domaines fonctionnels ou sont entièrement dupliquées pour donner des protéines 24TM avec des fonctions doubles (The major facilitator superfamily (MFS) revisited 2012).
Les TM1 et TM4 dans le faisceau N-terminal et les TM7 et TM10 dans le faisceau C-terminal, premier segment de chacune des répétitions de 3 TM, sont situés au centre du transporteur et forment :
la grande partie du site de liaison au substrat,
les régions d'occlusion.
Ces segments de TM subissent des changements conformationnels asymétriques lors de la liaison du substrat car elles possèdent une région non hélicoïdale.
Les TM7 et TM10 du faisceau TM C-terminal, liés par une pseudo-symétrie inversée, jouent un rôle critique.
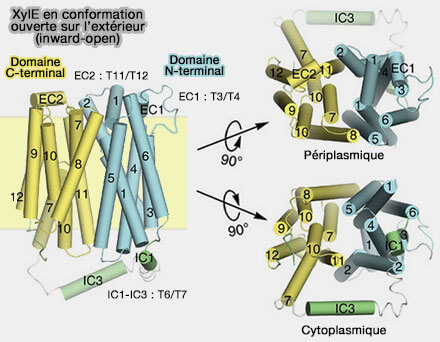
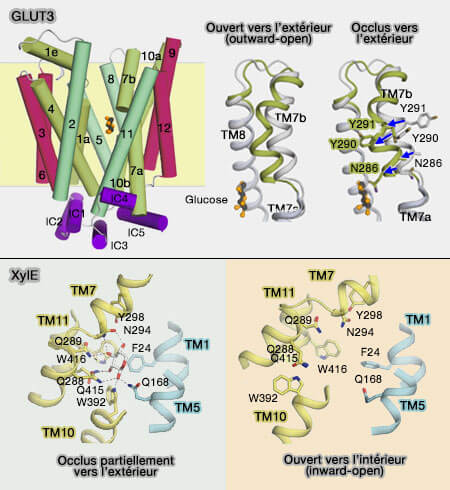
1. Dans GLUT3 (transporteur SLC2A3), uniport spécifique du transport du glucose dans les neurones, l'occlusion asymétrique vers l'extérieur est due à une flexion vers l'intérieur de la demi-hélice TM7b, située sur le faisceau C-terminal.
Les résidus de tyrosine volumineux voisins Tyr290 et Tyr291 sont responsables de l'occlusion induite par le substrat, bien qu'ils n'aient aucune interaction directe avec le sucre.
L'occlusion du site de liaison local d'un côté de la membrane est une condition préalable structurelle à la transition bascule-interrupteur qui ouvre l'accès au côté opposé (The SLC2 (GLUT) Family of Membrane Transporters 2014).
Le rôle central du substrat dans le mécanisme rocker-switch semble lié à la rupture et la reformation des ponts salins qui retiennent ensemble les faisceaux N et C-terminaux.
Dans les transporteurs de sucre, l'hélice TM10b fait partie d'un réseau de pont salin interfaisceau cytosolique strictement conservé. Quand TM7b se déplace vers l'intérieur pour coordonner le sucre, il n'interagit plus aussi étroitement avec le TM10, favorisant le mouvement de TM10b et la rupture du pont salin (Structure and mechanism of the mammalian fructose transporter GLUT5 2015).
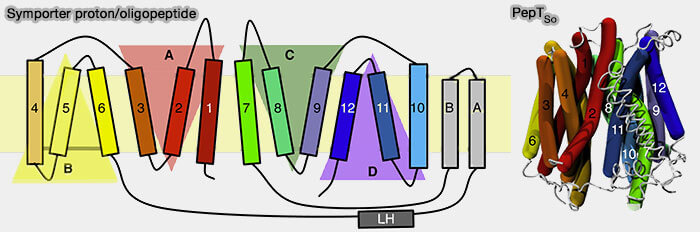
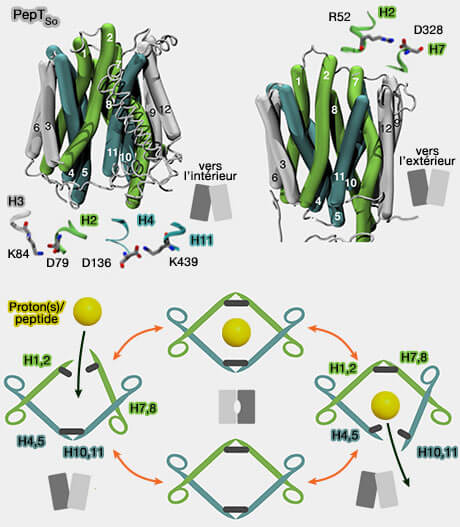
PepTSo : ponts salins et mécanisme
(Figure : vetopsy.fr d'après Folwer et coll)
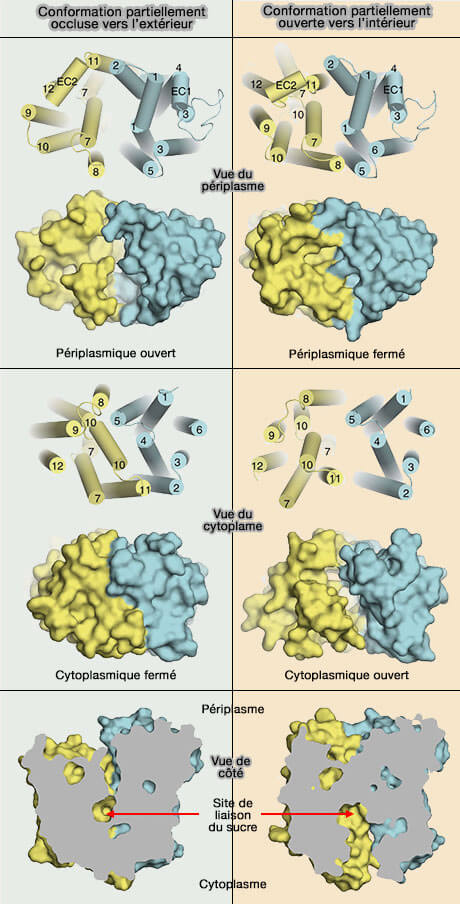
Dans les transporteurs GLUT, les ponts salins sont plus éloignés de la cavité centrale reliant les extrémités cytoplasmiques des TM3, TM4 et TM5 dans le faisceau transmembranaire N-terminal à celles des TM9, TM10 et TM11 dans le faisceau TM C-terminal (Molecular basis of ligand recognition and transport by glucose transporters 2015).
Ces résidus chargés, signature des transporteurs de sucre, sont retrouvés de la bactérie à l'homme. Les résidus chargés sont reliés par un axe de symétrie pseudo-double qui traverse le centre du transporteur et est perpendiculaire au plan de la membrane.
Les différents ponts salins peuvent être absolument nécessaires ou facultatifs selon les auteurs. Par exemple dans l'étude sur PepTSo, sept ponts salins ont été identifiés don les plus importants seraient (Gating Topology of the Proton-Coupled Oligopeptide Symporters 2015) :
D136-K439 (H4-H11) et K84-D79 (H3-H2), conservés dans la famille, qui stabiliseraient les conformations orientées vers l'extérieur,
R52-D328 (H2-H7), non conservés dans la famille SLC15, se formerait dans les conformations occluses et orientées vers l'intérieur.
La porte extramembranaire est formée par H1, H2, H7 et H8 et la porte cytoplasmique par H4, H5, H10 et H11 et semble être retrouvée dans toutes les 34 structures connues de MFS.
Les deux premières hélices de chacune des quatre unités répétées forment la moitié d'une des deux portes.
Ces structures renforcent le concept du mécanisme des transporteurs actifs lié à des répétitions inversées d'unités.