Quelles sont les structures mises en jeu et les phénomènes électriques intervenant dans les variations du potentiel membranaire ?
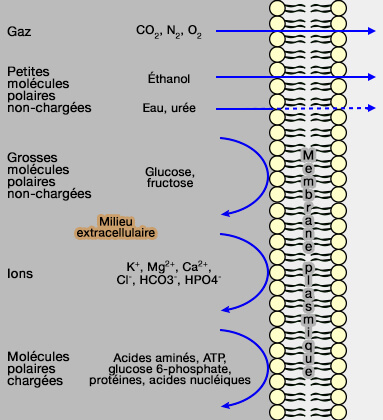
Perméabilité membranaire
(Figure : vetopsy.fr d'après Lodisch) au niveau de la membrane,
au niveau des milieux intracellulaire et extracellulaire.
Les potentiels membranaires sont créés par des différences de concentrations d'ions spécifiques de chaque côté de la membrane, et à la perméabilité sélective de cette membrane à certains ions, en particulier Na+, K+ Ca++et Cl-.
La mesure du potentiel de repos (entre -40 mV et -90 mV selon les neurones par exemple), montre que des ions sont transportés activement, i.e. contre leur gradient de concentration, à travers la membrane par des transporteurs actifs.
Des canaux ioniques sont responsables de la perméabilité sélective de la membrane à certains ions selon leur gradient de concentration.
Répartition
des concentrations ioniques
1. Le milieu intracellulaire contient en solution dans l'eau :
2. Le milieu extracellulaire ne renferme pas de grosses molécules en solution, mais des petites molécules et des ions.
Les milieux intracellulaire et extracellulaire sont de bons conducteurs électriques, contrairement à la membrane.
3. Si la membrane était perméable à tous les ions, les charges seraient réparties de chaque côté de la membrane et aucune différence de potentiel ne serait détectable.
Dans une cellule musculaire de mammifère (cf. tableau), le milieu intérieur est :
40 fois plus riche en ions K+,
12 fois moins riche en ions Na+ que le milieu extérieur.
La différence de potentiel est due à la distribution inégale des ions de part et d'autre de la membrane plasmique.
Ces concentrations ioniques différentielles sont constantes pour un type cellulaire donné ou un groupe donné.
Concentrations ioniques
des milieux intra et
extracellulaires
(mmoles/L)
Neurone de
Calmar
Neurone de
mammifère
Cellule musculaire
de mammifère
Intra
Extra
Intra
Extra
Intra
Extra
K+
400
20
140
5
155
4
Na+
50
440
5-15
145
12
145
Ca++
0,0001
10
0,0001
1-2
-
-
Cl-
40-150
560
4-30
110
4
1420
HCO3-
-
-
-
-
8
27
A-
-
-
-
-
155
-
Les cellules nerveuses et musculaires ont un milieu intérieur 20 à 100 fois plus riche en K+, 5 à 15 fois moins riche en Na+, 20 à 100 fois moins riche en Cl- que le milieu extérieur (autres concentrations ioniques).
Les ions K+ et Cl- sont répartis inversement les uns des autres. Par contre, il faut souligner que la plus grande partie des anions intracellulaires n'est pas constituée d'ions Cl- mais de gros anions protéiques (A-), qui ne peuvent traverser la membrane plasmique.
On voit que les animaux marins comme le Calmar ont des concentrations salines plus importantes que celles des mammifères.
Pour mieux comprendre, un exemple concret peut être utilisé (" Neurophysiologie" de R. F Schmidt - 1984 -). Soit une très petite portion de membrane séparant des volumes identiques de solutions salines différentes et contenant chacun 220 000 ions. On trouve :
100 000 ions K+ à l'intérieur de la cellule pour 2 000 à l'extérieur
10 000 ions Na+ à l'intérieur pour 108 000 à l'extérieur de la cellule.
La solution saline extracellulaire est essentiellement une solution de chlorure de sodium à 9 g/l, appelée solution saline physiologique car sa concentration en sels est à peu près identique à celle du sang.
Mouvements d'ions et signaux électriques
Théorie
La forte concentration intracellulaire des ions K+, qui peuvent passer librement la membrane par leurs canaux de fuite (nongated channel ou leakage channel), les force à quitter la cellule.
1. Cette pression osmotique, si rien ne s'y opposait, conduirait à une concentration identique en ions K+ de chaque côté de la membrane (lois de Fick, relation linéaire entre le flux de matière et son gradient de concentration).
Les deux lois de Fick, du nom du physiologiste allemand Adolf Eugene Fick (1829-1901) définissent la perméabilité membranaire ($P$) d'un soluté, en fonction du gradient de concentration ($\Delta C$) et du flux : $P=J/\Delta C$.
$P$ peut aussi s'exprimer en fonction de la solubilités membranaire ($\alpha$) en fonction du et du coefficient de diffusion ($D$) : $P=D\alpha/h$.
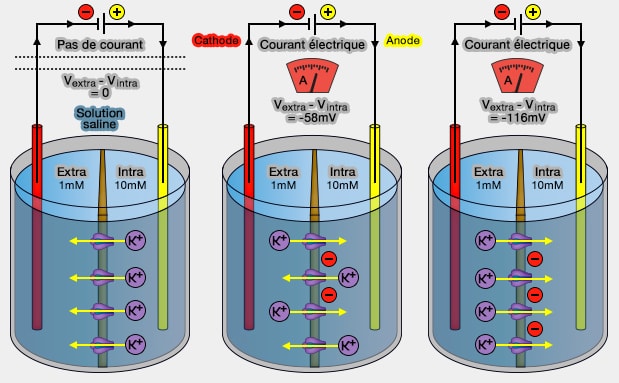
Flux ionique du potassium et potentiel de membrane
(Figure : vetopsy.fr)
2. Supposons que la membrane plasmique qui
sépare les deux milieux extra et intracellulaire soit semi-perméable, c'est-à-dire
qu'elle ne laisse traverser que les ions K, mais pas les ions ions Cl-.
Le passage d'un ion K+ d'un compartiment à l'autre conduit donc à l'apparition d'une charge positive dans le deuxième compartiment et d'une charge négative dans le premier, i.e. le Cl- qui y est resté.
À partir du passage d'un certain nombre d'ions K+, le gradient de potentiel établi par le flux lui-même, va s'opposer au passage des K+ (-58 mV dans la figure).
On peut alors définir un potentiel d'équilibre " théorique " de chaque ion, i.e. si la membrane était uniquement perméable à cet ion et s'il y avait alors un parfait équilibre entre :
le gradient de concentration qui pousse les ions K+ à sortir de la cellule en emportant les charges positives,
le gradient électrique qui tend à repousser les ions K+ et à les faire entrer dans la cellule.
$P_{Na}=1$, $P_{K}=40$ et $P_{Cl}=670$ : on trouve $V_m=-70,5\,mV$.
Dans les articles, on trouve différentes valeurs selon les cellules, ce qui est somme toute logique, mais aussi quelquefois selon les auteurs.
La création de la différence de potentiel, qui s'oppose à la diffusion, ne nécessite pas le passage d'un très grand nombre d'ions à travers la membrane, i.e. ces ne modifient pratiquement pas les concentrations
de départ des ions dans les deux compartiments (potentiel se repos).
Potentiels d'équilibre des ions incriminés
Quels sont les trois ions présents en quantité suffisamment significative (cf. tableau) ?
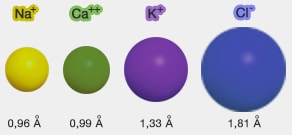
Ions et leurs rayons
(Figure : vetopsy.fr)
Ce sont les ions sodium (Na+), les ions potassium (K+) et les ions chlorure (Cl-).
La concentration des ions sodium et des ions chlorure est plus élevée dans le milieu extracellulaire.
La concentration des ions potassium est, au contraire, plus élevée dans le milieu intracellulaire.
On peut alors définir un potentiel d'équilibre " théorique " de chaque ion (cf. plus haut),
1. Le potentiel d'équilibre du potassium ($E_K$) serait la différence de potentiel mesurée si la membrane était uniquement perméable au potassium.
Les ions K+, dont la concentration est plus élevée dans le milieu intracellulaire que dans le milieu extracellulaire, sortiraient de la cellule.
La membrane cellulaire serait chargée négativement à sa face cytosolique et positivement sur sa face extérieure.
Or, c'est bien ce qui se passe car la membrane plasmique contient :
et des pompes qui accumulent K+ dans le cytoplasme,
et des canaux qui laissent passer les ions selon leur gradient de concentration.
Axone géant
de calmar
Motoneurone
de chat
$E_K$
-90 mV
-90 mV
$E_{Na}$
+55 mV
+60 mV
$E_{Cl}$
-50 mV
-70 mV
Le $E_K$ est proche de -90 mV.
2. Le potentiel d'équilibre du sodium ($E_{Na}$) serait la différence de potentiel mesurée si la membrane était uniquement perméable au sodium.
Les ions Na+, dont la concentration est plus élevée dans le milieu extracellulaire que dans le milieu intracellulaire, pénétreraient dans la cellule.
La membrane cellulaire serait chargée positivement à sa face cytosolique et négativement sur sa face extérieure.
Si nous reprenons l'exemple ci-dessus, pour une concentration de 10 mM de Na+ extracellulaire et 1 mM intracellulaire, on obtiendrait : $E_K=58log(10/1)=+58 mV$.
Cette valeur est " symbolique " car la membrane est considéré comme imperméable aux ions Na+, même si on a découvert récemment des canaux de fuite du sodium.
Dans l'axone géant de calmar (neurone de référence pour les expériences), $E_{Na}$, d'après la loi de Nerst serait de + 55 mV, + 60 mV dans le motoneurone du chat.
3. Le potentiel d'équilibre des chlorures ($E_{Cl}$) est la différence de potentiel mesurée si la membrane était uniquement perméable au sodium.
Ces potentiels ($E_{Na}$, $E_K$ et $E_{Cl}$) correspondent donc à des différences de potentiel théoriques entre l'intérieur et l'extérieur de la cellule.
Toutefois, on peut prédire que si, pour une raison quelconque, la membrane devient perméable à un de ces ions, le potentiel de membrane aura tendance à tendre vers une de ces valeurs.



{kind=link}

{kind=link}

 autres concentrations ioniques).
autres concentrations ioniques).