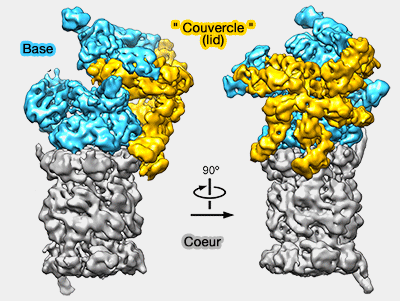
Le protéasome est une structure dynamique qui modifie sa conformation durant le processus et qui interagit avec un grand nombre de protéines.
La description ci-dessous concerne surtout la nomenclature, la structure, la conformation des différentes protéines : les positions décrites sont celles supposées du protéasome libre de tout substrat.
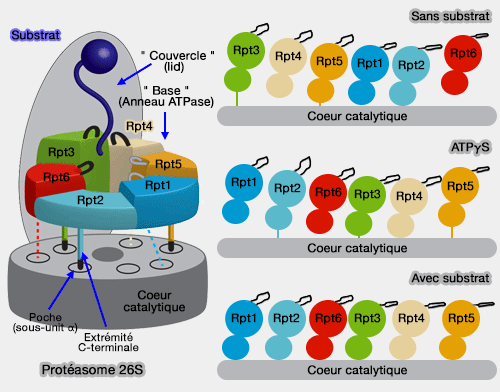
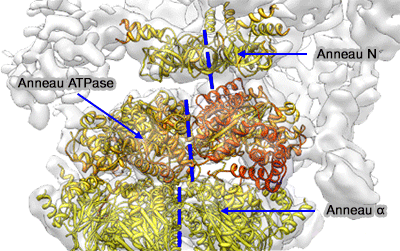
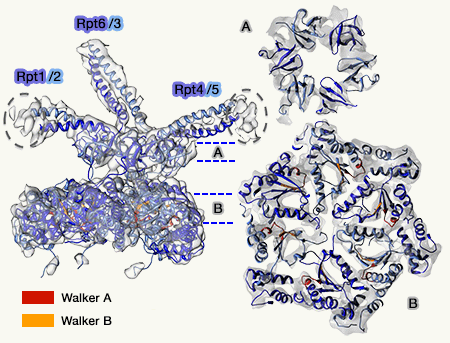
6 des 10 protéines, appelées Rpt1 à 6 (Regulatory particle triple-A protein ou Regulatory particle triphosphatase) forment un complexe héxamérique à activité ATPasique qui est directement en contact avec l'anneau α.
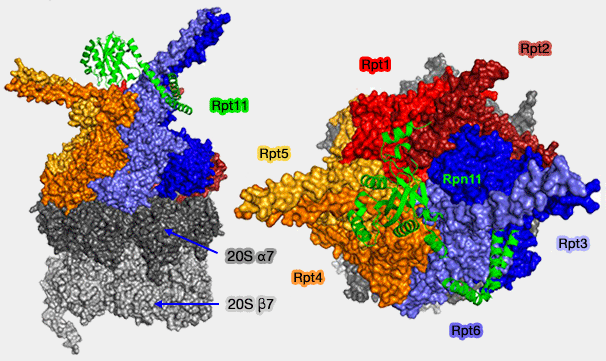
Anneau des Rpt avec Rpn11 (Rpn1, 2, 10 et 13 non représentés)
(Figure : vetopsy.fr d'après Yu)
Cet anneau constitue le moteur moléculaire du protéasome et est formé de trois modules dimériques : Rpt3-Rpt6, Rpt4-Rpt5 et Rpt1-Rpt2 (Structural Biology of the Proteasome 2013).
Structure des Rpt
Leurs structure est composée de deux domaines.
Domaine N-terminal
Le domaine N-terminal, est constitué par :
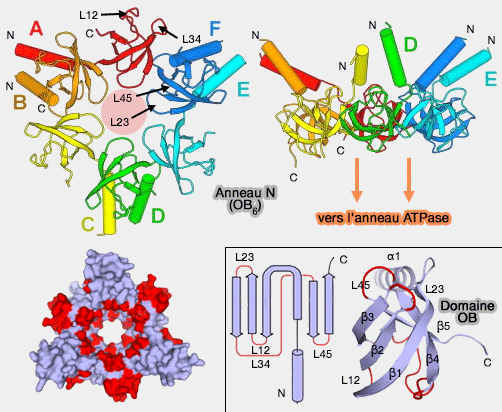
un domaine OB - (Oligonucleotide ou Oligosaccharide ou Oligomer Binding - (74-89 dans PAN),
Domaine OB et anneau OB ou N
(Figure : vetopsy.fr d'après Zhang)une hélice N-terminale (90-150 dans PAN).
Anneau OB
Les 6 domaines forment un anneau (anneau N ou OB), séparé de celui des ATPases, formé de 3 dimères : Rpt3/6, Rpt4/5 et Rpt1/2.
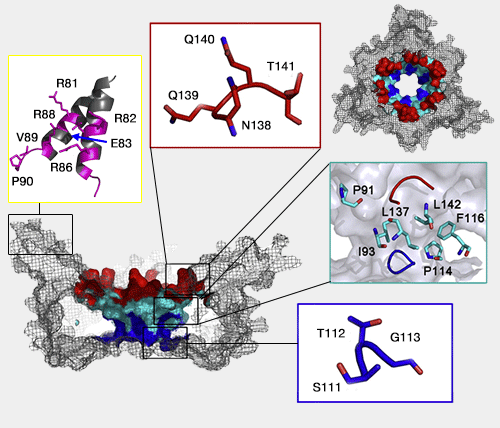
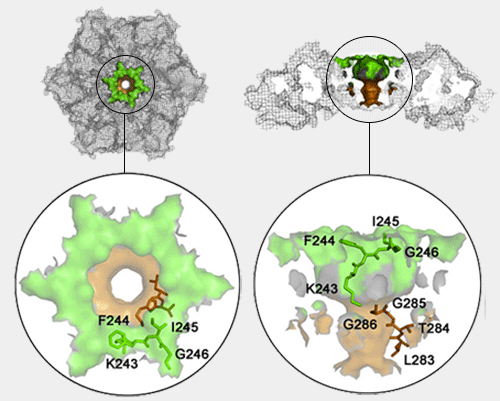
un anneau distal formé par la boucle L45 (Asn138-Gln139-Gln140-Thr141)
un anneau proximal, près de l'anneau ATPase, par la boucle L23 (Ser111-Thr-112-Gly113)
une surface intérieure hydrophobe entre les deux anneaux (Pro91, Ile93, Pro114, Pro116, Leu137 et Leu142).
Cette structure étant rigide et l'anneau proximal (13Å) étant bien plus étroit que l'anneau distal, le dépliage du substrat est probablement le résultat d'une translocation forcée à travers cet anneau.
Gly113, situé à la pointe de l'anneau proximal (L23), ne possède pas de chaîne latérale et permet une grande flexibilité de la chaîne principale. Sa substitution provoque une perte du dépliage et de la dégradation du substrat. Les substitutions sur l'anneau distal ont beaucoup moins d'effet.
Domaine OB et résidus impliqués
(Figure : vetopsy.fr d'après Zhang)
La surface hydrophobe joue aussi un rôle dans la facilitation de dépliage et la translocation du substrat : la substitution de Pro114 diminue fortement (70 à 80%) ce processus.
L'anneau N est faiblement connecté à l'anneau ATPase par une séquence flexible de 8 résidus - 150-158 - (cf. structure primaire) et l'axe et leur pore central ne coïncide pas.
Asp153 est présent chez PAN et les Rpt. Ala156 de PAN est remplacée par un acide aminé hydrophobe dans toutes les sous-unités Rpt. Ces caractéristiques sont susceptibles d'être fonctionnellement pertinentes.
Des substitutions sur ces résidus inhibent totalement le dépliage du substrat et diminue drastiquement sa dégradation, montrant bien que les deux anneaux (N et ATPase) coopèrent.
Le couplage entre les subcomplexes I et II implique des interactions transitoires par six acides aminés à la surface distale du subcomplexe II : Ser250, Leu251, Lys253, Asp254, Glu261 et Lys262 situés sur l'hélice α5 et sont conservés dans les sous-unités Rpt et doivent aussi jouer un rôle.
Superhélices (coiled coil) des Rpt (Rpt3/6, Rpt4/5, et Rpt1/2)
Les deux hélices N-terminales de chaque dimère (1/2, 3/6 et 4/5) forment trois domaines coiled-coil (superhélices) qui font protrusion et forment une sorte de tripode.
La superhélice de Rpt1/2 (27 résidus) semble interagir, car sa résolution est plus difficile, avec Rpn1 de la même façon que Rpt6/3 avec Rpn2.
La superhélice Rpt4/5 (33 résidus) n'entre pas en interaction avec les non-ATPases, mais elle semble montrer une grande flexibilité, ce qui fait que sa résolution est elle aussi difficile.
Les superhélices, par contre, contiennent un certain nombre de résidus polaires et chargés (R81/82/86/88, E83, V89, P9 chez PAN - voir les correspondances avec les Rpt dans l'article -), qui pointent à l'intérieur de l'entonnoir.
Étonnement, leurs substitutions par l'alanine, provoquent un accroissement du dépliage et de la dégradation du substrat, peut-être en diminuant leur action sur des résidus chargés négativement du substrat qui ralentissent le processus.
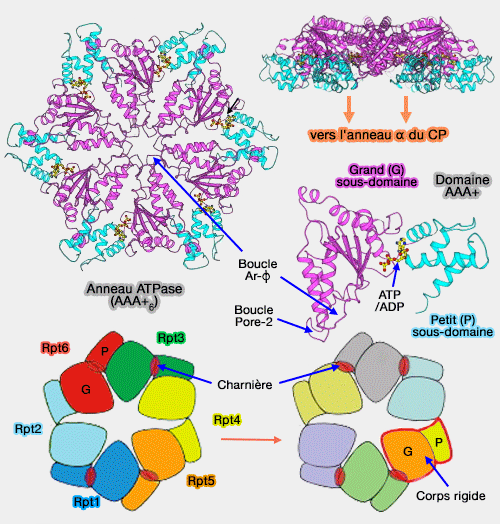
Les grands domaines des domaine AAA+ contiennent deux boucles essentielles aux fonctionnement du protéasome (pore loops) qui forment aussi deux anneaux comme pour les domaines OB.
Domaines AAA+ et résidus impliqués
(Figure : vetopsy.fr d'après Zhang)
1. L'anneau distal comprend la boucle Ar-Φ (Aromatic-aliphatic ou pore-1 loop) - Y/F-I/L/V-G -, leur permet de se lier à la protéine, de la déplier et de la transloquer au coeur catalytique.
Les extrémités C-terminales des Rpt permettent la fixation de la base à l'anneau α et lui permettent de suivre les modifications conformationnelles des Rpt dans les différentes phases du fonctionnement du protéasome.





{kind=link}
{kind=link}
{kind=link}