La sumoylation est une modification post-traductionnelle qui aboutit à la fixation covalente d'une (ou de plusieurs protéines SUMO) sur la protéine substrat.
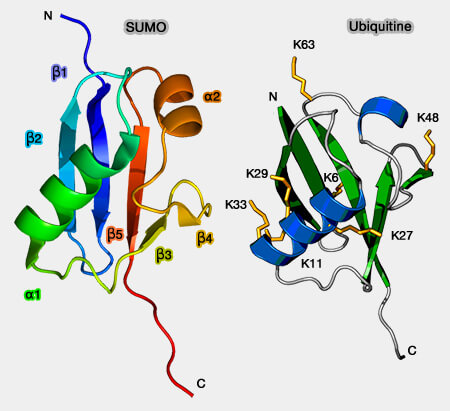
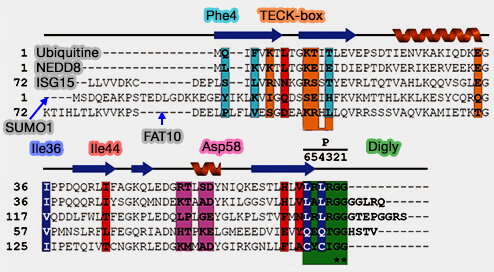
Les SUMO, comme d'autres UBL, diffèrent énormément de l'ubiquitine, quant à leur structure primaire : 20% seulement d'homologie.
Toutefois, ils possèdent une di-glycine C-terminale, attaché à un propeptide (variant suivant les SUMO : HSTV pour 1, VY pour 2, 12 aa pour 3).
Contrairement à l'ubiquitine, on connaît plusieurs isoformes : SUMO1, 2 et 3 (SUMO4 est décrite chez l'homme dans le rein, la rate et les noeuds lymphoïdes). Un nouveau SUMO, SUMO-5 a été trouvé en 2016 dans le PML bodies
Sumo2 et 3 sont presque identiques (97%) et comportent une extrémité N-terminale flexible (absente chez SUMO1, homologie à 50% avec les autres SUMO, et chez l'ubiquitine) qui leur permet de former des chaînes.
Séquences de l'ubiquitine et de différentes UBL
(Figure : vetopsy.fr d'après Ronau)
Les SUMO peuvent se lier à des nombreuse protéines (cf tables).
Toutes ces particularités expliquent leurs rôles différents de ceux de l'ubiquitine.
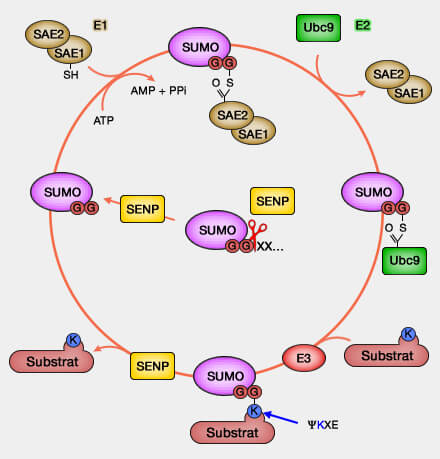
Mécanisme de
la sumoylation
Contrairement à l'ubiquitination qui conduit, en général, la protéine vers sa dégradation par le protéasome 26S, la sumoylation conduit à la stabilisation des complexes.
Leur rôle principal est de cibler certaines protéines impliquées dans la régulation de la transcription (SUMO: getting it on 2007), mais aussi dans la réparation de l'ADN, le transport nucléaire et cytosolique, l'apoptose, la stabilité des protéines, la réponse au stress et le cycle cellulaire (SUMO : a history of modification 2005).
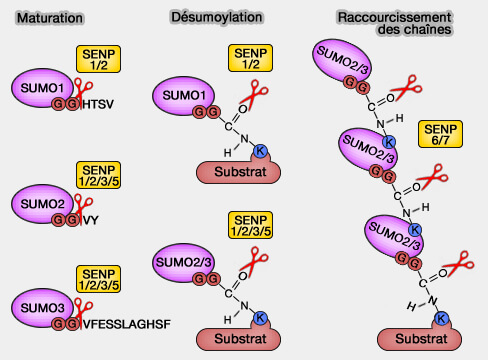
SENP chez l'Homme
(Figure : vetopsy.fr)
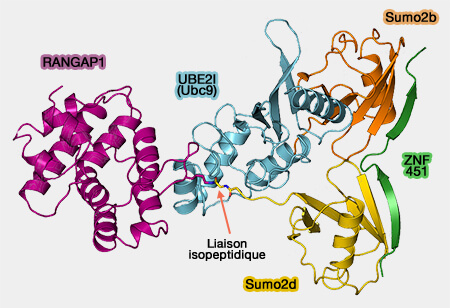
Par exemple, RanGAP1 (Ran GTPase-activating protein) est cytosolique alors que sumoylatée, elle se localise aux pores nucléaires par l'interaction avec la nucléoporine RanBP2 (NUP358).
SUMO, comme l'ubiquitine doit, pour se fixer sur la protéine, avoir recours à plusieurs enzymes.
SENP (sentrin- ou SUMO-
specific proteases)
La famille SENP (sentrin- ou SUMO-specific proteases) comprend 6 isoformes (SENP1, SENP2, SENP3, SENP5, SENP6 et SENP7) qui sont des hydrolases. Elles ont deux rôles spécifiques :
Elles clivent le propeptide des SUMO pour leur maturation.
Les SENP sont localisées dans différentes régions cellulaires.
SENP1 et SENP2 sont situées sur la membrane nucléaire et dans le nucléoplasme : leur rôle est d'inactiver les facteurs de transcription par sumoylation et leur activation par désumoylation, en particulier dans les processus de développement, en particulier par l'action de SENP2 sur PRC1 (Polycomb Repressive Complex 1), qui répriment de nombreux gènes précoces dont GATA…
SENP3 et SENP5 se trouvent dans le nucléole et sont impliquées dans la maturation précoces des ribosomes. On les retrouve aussi en plus faible proportion dans le nucléoplasme et même, dans le cytoplasme.
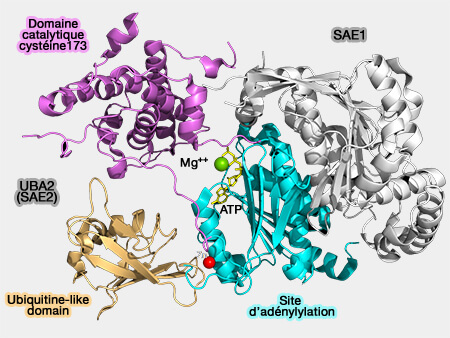
1. SUMO se lie à un enzyme E1 hétérodimère SAE (SUMO Activating Enzyme qui comprend SAE1 et SAE2 - appelé aussi UBA2, Ubiquitin-like 1-activating enzyme E1B).
Ube2I se distingue des autres enzymes de conjugaison par sa capacité à reconnaître directement les protéines visées.
Ube2I-SUMO peut donc catalyser la formation d'une liaison isopeptidique entre le groupe carboxyle ($\ce{-C(=O)OH}$) C-terminal de SUMO et la lysine (K) de la protéine, à condition que la lysine soit située dans un motif SUMO de conjugaison.
Le motif consensus pour l'attachement de SUMO est un tétrapeptide Ψ-K-x-D/E ou Ψ représente un acide aminé hydrophobe, K la lysine acceptrice qui forme la liaison isopeptidique avec SUMO, x un acide aminé quelconque et D/E représente un acide aspartique ou un acide glutamique. Cependant d'autres facteurs semblent intervenir.
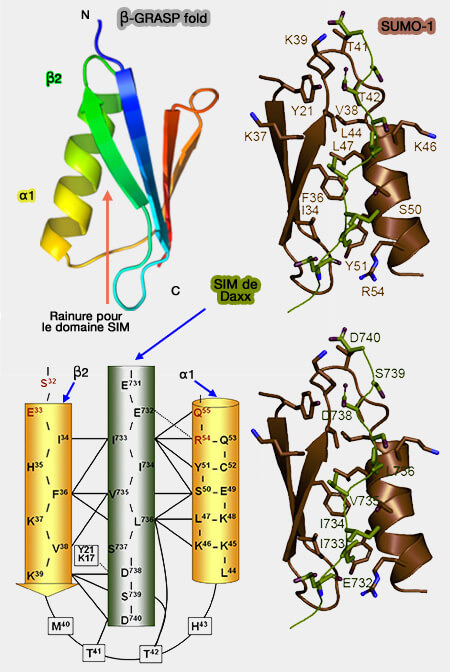
SIM de Daxx et β-GRASP fold de SUMO-1
(Figure : vetopsy.fr d'après Chang)
Le site actif de Ube2I contient un motif CES/D : Conserved E2 Serine/Aspartate.
Cette sumoylation peut se faire directement sans E3 ligases.
E3 ligases éventuelles
Un petit nombre de ligases E3 peuvent accélérer ce phénomène selon différentes interactions.